
Monday, June 08, 2009
|
|
37th Common
|
|
Join Date: Mar 2006
Posts: 1,514
Thanks: 1,053
Thanked 1,681 Times in 873 Posts
|
|
 Cell Biology Golgi Apparatus
Cell Biology Golgi Apparatus
Golgi Apparatus
The Golgi apparatus (GA), also called Golgi body or Golgi complex and found universally in both plant and animal cells, is typically comprised of a series of five to eight cup-shaped, membrane-covered sacs called cisternae that look something like a stack of deflated balloons. In some unicellular flagellates, however, as many as 60 cisternae may combine to make up the Golgi apparatus. Similarly, the number of Golgi bodies in a cell varies according to its function. Animal cells generally contain between ten and twenty Golgi stacks per cell, which are linked into a single complex by tubular connections between cisternae. This complex is usually located close to the cell nucleus.
Discovery
Due to its relatively large size, the Golgi apparatus was one of the first organelles ever observed. In 1897, an Italian physician named Camillo Golgi, who was investigating the nervous system by using a new staining technique he developed (and which is still sometimes used today; known as Golgi staining or Golgi impregnation), observed in a sample under his light microscope a cellular structure that he termed the internal reticular apparatus. Soon after he publicly announced his discovery in 1898, the structure was named after him, becoming universally known as the Golgi apparatus. Yet, many scientists did not believe that what Golgi observed was a real organelle present in the cell and instead argued that the apparent body was a visual distortion caused by staining. The invention of the electron microscope in the twentieth century finally confirmed that the Golgi apparatus is a cellular organelle.
Structure
In general, the Golgi apparatus is made up of approximately four to eight cisternae, although in some single-celled organisms it may consist of as many as 60 cisternae. Matrix proteins hold the cisternae together, and the whole of the Golgi apparatus is supported by cytoplasmic microtubules. The apparatus has three primary compartments, known generally as cis (cisternae nearest the endoplasmic reticulum), medial (central layers of cisternae), and trans (cisternae farthest from the endoplasmic reticulum). Two networks, the cis Golgi network and the trans Golgi network, which are made up of the outermost cisternae at the cis and trans faces, are responsible for the essential task of sorting proteins and lipids that are received (at the cis face) or released (at the trans face) by the organelle.
Transport of substances
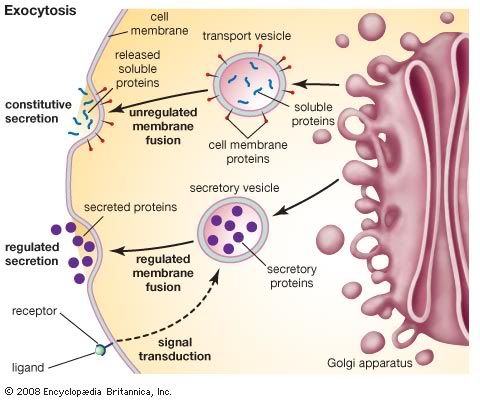
The proteins and lipids received at the cis face arrive in clusters of fused vesicles. These fused vesicles migrate along microtubules through a special trafficking compartment, called the vesicular-tubular cluster that lies between the endoplasmic reticulum and the Golgi apparatus. When a vesicle cluster fuses with the cis membrane, the contents are delivered into the lumen of the cis face cisterna. As proteins and lipids progress from the cis face to the trans face, they are modified into functional molecules and are marked for delivery to specific intracellular or extra cellular locations. Some modifications involve cleavage of oligosaccharide side chains followed by attachment of different sugar moieties in place of the side chain. Other modifications may involve the addition of fatty acids or phosphate groups (phosphorylation) or the removal of monosaccharides. The different enzyme driven modification reactions are specific to the compartments of the Golgi apparatus. For example, the removal of mannose moieties occurs primarily in the cis and medial cisternae, whereas the addition of galactose or sulfate occurs primarily in the trans cisternae. In the final stage of transport through the Golgi apparatus, modified proteins and lipids are sorted in the trans Golgi network and are packaged into vesicles at the trans face. These vesicles then deliver the molecules to their target destinations, such as lysosomes or the cell membrane. Some molecules, including certain soluble proteins and secretory proteins, are carried in vesicles to the cell membrane for exocytosis (release into the extracellular environment). The exocytosis of secretory proteins may be regulated, whereby a ligand must bind to a receptor to trigger vesicle fusion and protein secretion.

|