CSS Forums
Saturday, April 20, 2024
01:16 AM (GMT +5)
01:16 AM (GMT +5)





|
#81
 Sunday, May 17, 2009
Sunday, May 17, 2009
|
|||
|
|||
|
SKIN The skin is the outer covering of the body. It is the largest organ of the integumentary system made up of multiple layers of epithelialtissues, and guards the underlying muscles, bones, ligaments and internal organs.[1] The adjective cutaneous literally means "of the skin" (from Latin cutis, skin). Because it interfaces with the environment, skin plays a very important role in protecting (the body) against pathogens. Its other functions are insulation, temperature regulation, sensation, synthesis of vitamin D, and the protection of vitamin B folates. Severely damaged skin will try to heal by forming scar tissue. This is often discolored and depigmented. In humans, skin pigmentation varies among populations, and skin type can range from dry to oily. Skin performs the following functions: · Protection: an anatomical barrier from pathogens and damage between the internal and external environment in bodily defense; Langerhans cells in the skin are part of the adaptive immune system. · Sensation: contains a variety of nerve endings that react to heat and cold, touch, pressure, vibration, and tissue injury; see somatosensory system and haptics. · Heat regulation: the skin contains a blood supply far greater than its requirements which allows precise control of energy loss by radiation, convection and conduction. Dilated blood vessels increase perfusion and heat loss while constricted vessels greatly reduce cutaneous blood flow and conserve heat. Erector pili muscles are significant in animals. · Control of evaporation: the skin provides a relatively dry and impermeable barrier to fluid loss. Loss of this function contributes to the massive fluid loss in burns. · Aesthetics and communication: others see our skin and can assess our mood, physical state and attractiveness. · Storage and synthesis: acts as a storage center for lipids and water, as well as a means of synthesis of vitamin D by action of UV on certain parts of the skin. · Excretion: sweat contains urea, however its concentration is 1/130th that of urine, hence excretion by sweating is at most a secondary function to temperature regulation. · Absorption: Oxygen, nitrogen and carbon dioxide can diffuse into the epidermis in small amounts, some animals using their skin for their sole respiration organ. In addition, medicine can be administered through the skin, by ointments or by means of adhesive patch, such as the nicotine patch or iontophoresis. The skin is an important site of transport in many other organisms. · Water resistance: The skin acts as a water resistant barrier so essential nutrients aren't washed out of the body skin functions The skin not only gives us our appearance and shape, it also serves other important functions: Protection from · mechanical impact (i.e. pressure, stroke) · thermic impact (i.e. heat, cold) · chemical impact (i.e. acids) · microorganisms (bacteria, viruses, fungi) · UV-radiation · water loss Immune function Besides providing a biological barrier against microorganisms through its acidic pH-value, the skin is immunologically active through defense mechanisms in epidermis and dermis. Temperature regulation Through sweat-producing glands and the evaporation of sweat and water, the body temperature is controlled. Another mechanism for rapid cooling is vasodilation (widening of blood vessels). Through vasoconstriction (narrowing of blood vessels), heat loss is prevented. Sensation Through nerve endings and receptors in the skin, sensations such as touch, pain, heat or cold are processed. Vitamin production The skin produces Vitamin D through exposure to ultraviolet radiation in sunlight. Social-interactive Through paling, blushing and other expressions regulated by the autonomic nervous system, the skin serves as a communication system. All vertebrate bodies are covered with skin. The skin may be smooth, rough, slimy, or scaly, but it's still skin. Vertebrate skin has a covering of hair, feathers, or scales. There are about 45,000 different kinds of vertebrates living on Earth. Scientists have classified vertebrates into five smaller groups, based on characteristics they share. These groups are the mammals, birds, reptiles, amphibians, and fish. 
|






|
#82
Sunday, May 17, 2009
|
|||
|
|||
|
BONE What types of bone are found in vertebrates? Vertebrates possess two types of bone. Dermal bone consists of bony structures (plates and scales) that develop in the skin. The bony armor of the earliest jawless fish was dermal bone; so are shark scales, shoulder blades, and the roof of your skull. Dermal bone does not form from cartilage first and then calcify, but endoskeletal bone does; in fact, in cartilaginous fishes it may never form true bone. Vertebrae, ribs, appendages, and the jaw are endoskeletal bone. The vertebrate skull is actually a complex structure of both endoskeletal and dermal bone. All vertebrates have cartilage in addition to bone, or instead of bone. Cartilage may be flexible, like the cartilage in your nose and ears, or hard and firm, like the cartilage you can feel in your larynx (voicebox). Cartilage also covers the adjoining surfaces of bones in movable joints; damage or breakdown of this cartilage results in painful movement. The calcified cartilage that makes up shark teeth and vertebrae is not true bone; it is dead when functional, whereas bone is a living tissue. Bone Anatomy Bone (anatomy), hard connective tissue, the major component of almost all skeletal systems in adult vertebrate animals. Bone appears to be nonlivingin fact, the word skeleton is derived from a Greek word meaning dried up. However, bone actually is a dynamic structure composed of both living tissues, such as bone cells, fat cells, and blood vessels, and nonliving materials, including water and minerals. Bones are multipurpose structures that play diverse, vital roles in vertebrates. They provide a framework for the body, supporting it and giving it shape. They also provide a surface for the attachment of muscles and act as levers, permitting many complex movements. Many bones protect softer internal organs; for example, skull bones protect the brain, and rib bones form a cage around the lungs and heart. In addition to these structural and mechanical functions, bones also participate in the bodys physiology. They store calcium, a mineral essential for the activity of nerve and muscle cells. The soft core of bone, thebone marrow, is the site of formation of red blood cells, certain white blood cells, and blood platelets. An adult human has 206 bones, which account for 14 percent of the bodys total weight. The longest and strongest bone is the thighbone, which at maturity is about 50 cm (20 in) long and 2.5 cm (1 in) wide. The smallest bone, the stirrup bone, is one of three tiny bones buried within the middle ear; it is only 0.18 cm (0.07 in) long. Bone Structure and Composition Bone consists of living cells widely scattered within a nonliving material called the matrix. The matrix is formed by osteoblasts, cells that are constantly renewed in the bone. Osteoblasts make and secrete the protein collagen, which makes bones elastic so that they can give under the stresses generated by walking, lifting, and other activities. Osteoblasts also secrete mineral salts formed from calcium and phosphorus, which impart hardness so that bones do not break easily. If more bone is needed, new osteoblasts carry out the task of building it. As bone tissue matures, osteoblasts transform into osteocytes, mature bone cells that carry out daily cellular activities. There are two main types of bone. Compact bone, which makes up most of the bone of arms and legs, is very dense and hard on the outside. The structural units of compact bone are osteons, elongated cylinders that act as weight-bearing pillars, able to withstand any mechanical stress placed on the bone. The center of each osteon contains a hollow canal that acts as a central passageway for blood vessels and nerves. In some bones, internal to the compact bone is spongy bone, also known as cancellous bone, composed of a honeycomb network of bones called trabeculae that act as supporting beams. Spongy bone is designed to bear stress from several directions, such as that exerted on the pelvis in bending or stretching. The spaces between the trabeculae are filled with red bone marrow containing the blood vessels that nourish spongy bone. Spongy bone is found in bones of the pelvis, ribs, breastbone, vertebrae, skull, and at the ends of the arm and leg bones. Surrounding both compact and spongy bone is a thin membrane, the periosteum. The outer layer of this membrane contains nerves and blood vessels that branch and travel into the bone. The inner layer of the periosteum consists mainly of osteoblasts. The point where two or more bones come together is called a joint, or articulation. Different kinds of joints enable different ranges of motion. Some joints barely move, such as those between the interlocking bones of the skull. Other bones, held together by tough connective tissues called ligaments, form joints such as the hinge joint in the elbow, which permits movement in only one direction. The pivot joint between the first and second vertebrae allows the head to turn from side to side. Intimately associated with bone is another type of connective tissue called cartilage. Cartilage is softer, more elastic, and more compressible than bone. It is found in body parts that require both stiffness and flexibility, such as the ends of bones, the tip of the nose, and the outer part of the ear. Development and Maintenance During the early development of a baby within its mothers body, the skeletal structure consists of cartilage. At about the eighth week of fetal development, calcium and phosphorus salts begin to deposit around the cartilage. At 40 weeks of development, however, the fetal bones still consist primarily of soft cartilage. The skull consists of several cartilage plates that are not completely joined. The spaces between the cartilage plates are called soft spots, or fontanels. The soft cartilage and the fontanels enable the skull to be compressed as it passes through the birth canal. During childhood, cartilage gradually is replaced by bone through the activity of osteoblasts. More than 300 bones are present in an infant, several of which fuse as the infant matures. Throughout life, bone tissue undergoes continual breakdown and restoration in response to the bodys demands. For example, calcium must always be present in blood at a certain concentration. If calcium blood levels drop, cells called osteoclasts break down bone to release calcium into the bloodstream. If exercise increases muscle mass, bones must thicken so that the pull of the stronger muscle does not break the bone. In this case, osteoblasts create new bone. During childhood and adolescence, much more bone tissue is deposited than broken down, so the skeleton grows in size and strength. During early adulthood, breakdown slowly begins to exceed deposits. As a person ages, bone tissue is depleted, and bones are weakened and increasingly susceptible to breaks. Exercise and proper diet are critical for maintaining healthy bone growth at all stages of life. Nutrientsparticularly sufficient calcium, phosphorus, and vitamin D, and hormonesincludinggrowth hormone, parathyroid hormone, calcitonin, and sex hormonesall influence bone growth. Fractures, or breaks, are very common injuries to bones. The repair process requires the interplay of several processes. About a week after a fracture occurs, cells from the periosteum invade the damaged area and produce a fibrous network. Then, other cells produce cartilage in the network. Finally, osteoblasts enter the network and convert the cartilage to bone. Complete healing may take weeks or even months, depending on the individuals general health, age, and other factors. Some fractures are treated with a splint, a firm object that supports the area surrounding the broken bone and restricts motion. Other fractures must be completely immobilized to heal because movement can cause a new fracture in the area. These fractures may be immobilized with a cast, plastic or plaster wrapped around the area that surrounds the broken bone. 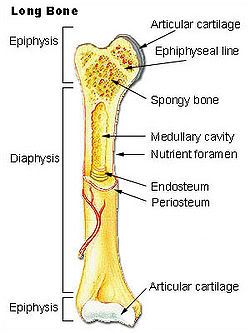 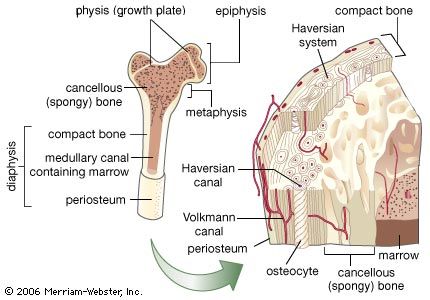 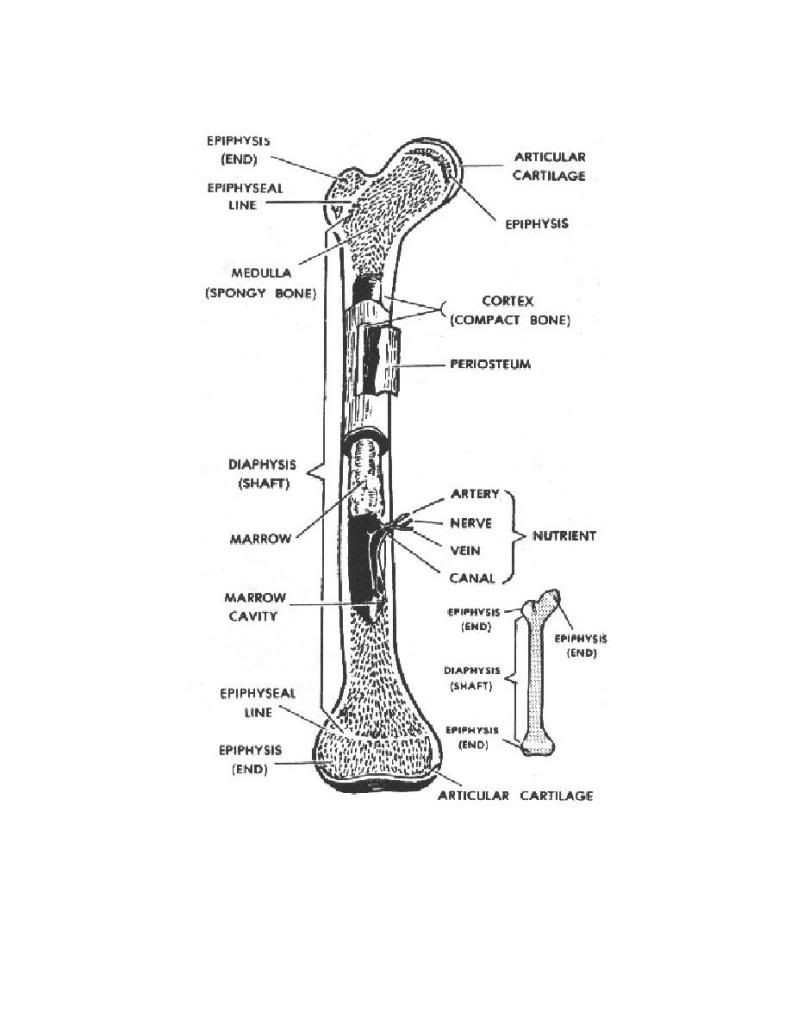
|
|
#83
Monday, May 18, 2009
|
|||
|
|||
|
AMPHIOXUS, or Lancelet, the name of small, fish-like, marine creatures, forming the class Cephalochorda, of the phylum Vertebrata.
Amphioxi are small marine animals found widely in the coastal waters of the warmer parts of the world and less commonly in temperate waters. Both morphological and molecular evidence show them to be close relatives of the vertebrates. Amphioxi are seldom more than 8 cm (3 inches) long and resemble small, slender fishes without eyes or definite heads. They are grouped in two generaBranchiostoma (also called Amphioxus) and Epigonichthyes (also called Asymmetron)with about two dozen species. The chordate featuresthe notochord (or stiffening rod), gill slits, and dorsal nerve cordappear in the larvae and persist into adulthood. Amphioxi spend much of their time buried in gravel or mud on the ocean bottom, although they are able to swim. When feeding, they let the anterior part of the body project from the surface of the gravel so that they can filter food particles from water passing through their gill slits. At night they often swim near the bottom. They burrow into sand using rapid movements of the body, which is tapered at both ends and is covered by a sheath (the cuticle). The animals swim by contracting the muscle blocks, or myotomes, that run from end to end on each side of the body. The blocks on each side are staggered, producing a side-to-side movement of the body when swimming. Amphioxi are not buoyant, and they sink quickly when they stop swimming. A dorsal fin runs along the entire back, becomes a caudal fin around the tip of the tail, and then continues as a ventral fin; there are no paired fins. The notochord runs through the body from tip to tip, providing a central support. A slight bulge distinguishes the anterior end of the nerve cord. Although there is no brain or cranium, growing evidence suggests that the vertebrate brain evolved from a portion of the nerve cord in a lancetlike ancestor. Blood flows forward along the ventral side and backward along the dorsal side, but there is no distinct heart. The oral cavity of amphioxi is furnished with a hood whose edges are lined with cirri; these are fringelike structures that form a coarse filter to screen out particles too large to be consumed. Water is directed through the small mouth into the pharynx by the action of cilia on the gill slits. Food particles in the passing water are caught by the mucous lining of the gill basket and passed into the gut, where they are exposed to the action of enzymes. Unlike other chordates, amphioxi are capable of a digestive process called phagocytosis, in which food particles are enveloped by individual cells. Above the pharynx is the excretory system made up of the nephridia, which opens into an excretory canal leading to the atrium. The endostyle corresponds to the thyroid in vertebrates, since it seems to produce iodinated, tyrosine molecules, which may function as regulatory substances, much like hormones, in amphioxi. Male and female amphioxi are identical in outward appearance and differ internally only in the nature of the gonads, or reproductive glands, which form in rows on the wall of the atrial cavity. Breeding takes place several times a year in tropical regions but only once in temperate areas. Sacs containing eggs or sperm burst and discharge their contents into the water through an opening on the underside of the body. Eggs are fertilized in the water, and after about two days microscopic ciliated larvae develop from the fertilized eggs. The larvae are carried with ocean currents for several weeks before metamorphosing into juvenile amphioxi and taking up life in sandy sediments. Along parts of the coast of China, amphioxi are so numerous that they constitute the basis of a fishing industry. What appear to be fossilized amphioxi have been found recently in sediments about 525 million years old. 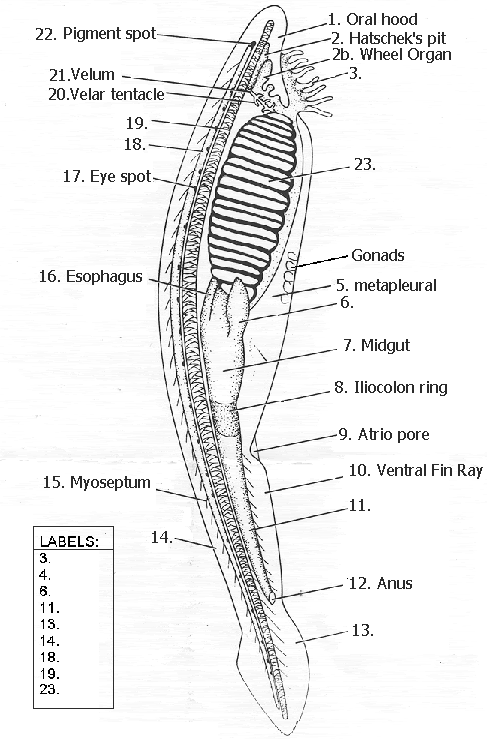 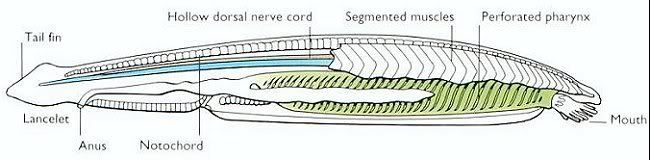
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#84
Monday, May 18, 2009
|
|||
|
|||
|
Amphibian heartFrogs are known for their three-chambered heart, which they share with all tetrapods except birds and mammals. In the three-chambered heart, oxygenated blood from the lungs and de-oxygenated blood from the respiring tissues enter by separate atria, and are directed via a spiral valve to the appropriate vesselaorta for oxygenated blood and pulmonary vein for deoxygenated blood. This special structure is essential to keeping the mixing of the two types of blood to a minimum, which enables frogs to have higher metabolic rates, and to be more active than otherwise.
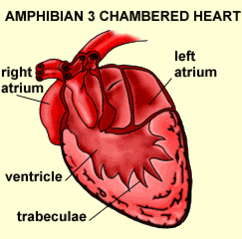 Amphibian circulation has a more complex arrangement of blood vessels, forming a double-circuit circulation. The three chambered amphibian heart consists of the right and left atria and a single ventricle. In the amphibian heart, oxygenated and deoxygenated blood are kept somewhat separate, largely as a result of the timing of the contractions of the left and right atria. First the blood returning from the right atrium is pumped by the ventricle into pulmonary circulation. After most of the deoxygenated blood has left the ventricle, oxygenated blood is pushed in and pumped through the amphibian body. 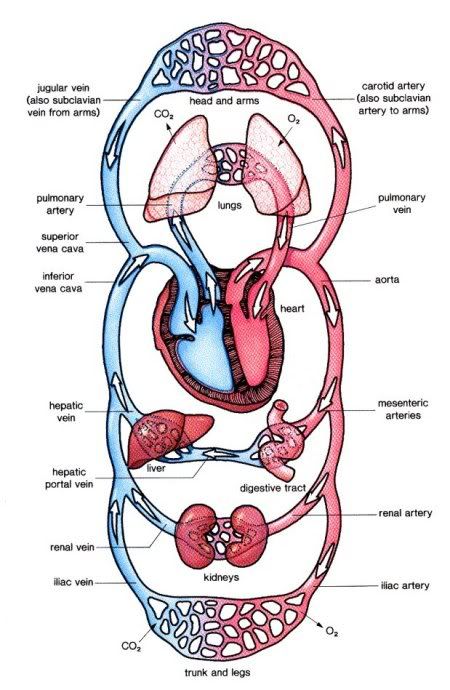
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#85
Tuesday, May 19, 2009
|
|||
|
|||
|
Aortic Arches in Vertebrates The circulatory system provides these benefits: (1) the transport of gases mainly O2 and CO2, (2) the transport of waste products from tissues to gills, lungs, kidneys, and skin, (3) the transport of nutrients from the intestines, (4) the transport of hormones, (5) the transport of agents of the immune system, (6) osmoregulation, and (7) thermoregulation. The first vertebrates solved many anatomical challenges simultaneously. These systems had to co-evolve because they are interdependent. Improvements in one system increased selection for improvements in one or more other systems. For example, the advent of a muscular pharynx led to the evolution of jaws and greatly accelerated efficiency in the rate of food acquisition. Concomitantly, the ability to pump large volumes of water through the pharynx allowed gas exchange organs (gills) to be concentrated there. Improvements in the respiratory and digestive systems demanded a means to distribute gases and nutrients to the body tissues (and the metabolic wastes away from the tissues). The heart developed as a specialized pump. We will look specifically at the heart in the next chapter. Here, we will review the major evolutionary changes in vertebrate arterial blood vessels in response to demands imposed by the respiratory system (advent of lungs) and the digestive system (the metabolic optimality required by homeothermy). Overall, the biggest shift in circulation pattern throughout the vertebrates is from a single loop to a double loop. A single loop is one that goes from the heart to the gills to the body and back to heart, as in most fishes. A double loop goes from the heart to the lungs and back (pulmonary circuit) and then from the heart to the body and back (systemic circuit). Lungfishes, amphibians, reptiles, birds and mammals show varying degrees of double loop circulation. Evolution of the cardiovascular system The early evolution of the cardiovascular system is tied closely to the early changes in the pharynx. Recall that the early pharynx of pre-vertebrates was a basket used for filter feeding. It later became vascularized and associated with gills with a large surface area for gas exchange. The ancestral pattern of blood flow was: body _ heart _ gills _ body. All fishes except the lung fishes use this pattern. The number of branchial arches bearing gills declined from the jawless fishes (Agnatha) to the jawed fishes (Gnathostoma), in part because one of the arches formed the margin of the newly evolved jaw, but also because of an overall reduction in pharyngeal arches. The jawless hagfish, representative of the simplest vertebrate class, has 15 aortic arches. Lamprey, representative of the next simplest vertebrate class, have 7 aortic arches. The first arch was lost somewhere between the jawless and the jawed fishes. The second arch formed the skeletal elements of the jaw margin, leaving 5 branchial arches bearing gills. 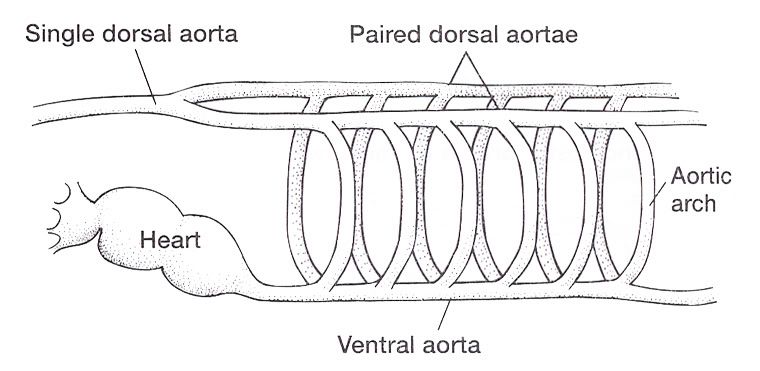 The arches bearing gills (and therefore supplied with blood) are numbered #2, #3, #4, #5, and #6, reflecting the ancestral pattern. This is the starting point for vessel evolution in air-breathing vertebrates. Advanced fishes, the teleosts, have since simplified their pharynx to only four pairs of gills on arches #3, 4, 5, and 6, but theyre on a line of evolution separate from the one that led to mammals. The ancestral pattern of blood flow is a conus arteriosus leading cranially from the single-chambered ventricle and forming a ventral aorta running along the base of the pharynx. Afferent branchial arteries (low O2) branch left and right at each branchial arch to supply blood to gills on each side of the body. Blood exits the gills dorsally via efferent branchial arteries (high O2). Efferent branchial arteries join with others from their side of the pharynx to form two short dorsal aortae that flow caudally. The two aortae converge at the caudal end of the pharynx to form a single dorsal aorta that continues caudally to supply oxygenated blood to the body tissues. Oxygen content of air is about 20%, which is many times greater than the saturation of dissolved oxygen in water. Water saturates at about 0.001% oxygen under ideal conditions. In warm temperatures, the ability of water to hold oxygen declines. Decay (=oxidation) of organic matter by microbial organisms commonly removes some or even all oxygen from the water resulting in fish mortality. Fishes with the ability to access atmospheric oxygen are favored in habitats where water oxygen levels frequently fall to close to zero. The fleshy-lobed-finned lungfishes developed an out pocketing of the esophagus that enabled them to swallow air and hold it for extended periods. Extraction of supplemental oxygen was enhanced by subsequent vascularization of the "lung". Blood supply came from the nearest blood vessel, which happened to be the efferent branchial artery of the 6th aortic arch. This vessel became the pulmonary artery and marks the beginning of a double-loop circulation pattern of blood flow. Blood returns from the lung to the atrium via a pulmonary vein and boosts the oxygen content of venous circulation. The atrium in lungfishes becomes partially divided to keep oxygenated blood from the lung separate from the deoxygenated blood returning from the body. The 6th aortic arch that supplies blood to the lung retains a connection to the pair of dorsal aortae. This connection is called the ductus arteriosus. 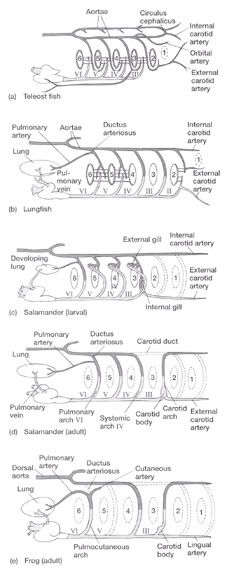 Amphibians Amphibians inherit the lungfish pattern of blood flow with a few modifications. The second aortic arch is lost from the aquatic phase of larval development. In larval salamanders, branchial arches #3-5 supply blood to the external gills. The 6th aortic arch supplies the developing lung. Frog tadpoles have internal gills supplied by aortic arches #3-6. In adult amphibians the external gills are lost but the aortic arches remain as systemic vessels. Arches #4, 5, 6 supply blood to the body (and lung and skin) caudal to the heart, while arch #3 supplies blood to the head region. The segment of the paired dorsal aortae between arches #3 and #4 is lost in adult amphibians effectively separating blood flowing to the head region from blood flowing to the rest of the body. This segment is called the carotid duct. Closure of the carotid duct boosts the pressure of blood going to the head. The ductus arteriosus is retained in adult salamanders but lost in adult frogs. The 5th aortic arch is also lost. Frogs therefore have completely separate pulmonary and systemic circuits of blood circulation. Arch #4 supplies the systemic circuit, arch #6 functions as the pulmonary artery, and arch #3 forms the carotid arteries. In lungless salamanders (plethodonts) aortic arch #6 supplies blood to the skin of the neck and back and thus still serves as the blood supply for gas exchange at the respiratory surface. Reptiles Reptiles retain left and right aortic arches #3, 4, and 6. Arch #5 is lost. Arch #3 forms the left and right carotid arteries leading to the head. Arch #4 forms the left and right aortic arches that supply the rest of the systemic circuit. The aortic arches continue to follow the path set by gills that no longer exist. The aortic arches head cranially from the heart to where the ancestral gills used to be, then loop caudally and dorsally on the left and right side, again, as if to supply gills on the left and right side of the pharynx. The aortic arches meet above the heart and converge to form a single dorsal aorta flowing caudally to supply the body tissues. Arch #6 forms the single pulmonary artery. In summary, the ventral aorta leading from the ventricle is subdivided into 3 vessels; the left and right systemic arches (arch #4), and the pulmonary trunk (arch #6). The right systemic arch is greater in size than the left systemic arch. The carotid duct joining the third and fourth aortic arches remains open in reptiles. Frogs are specialized amphibians much derived from the now-extinct anthracosaur amphibians that gave rise to the reptiles. 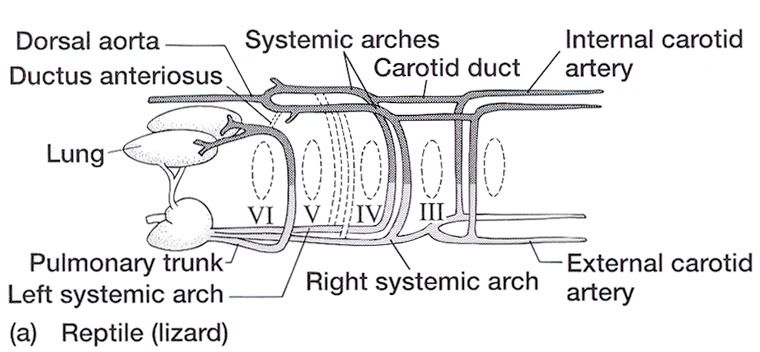 Birds Birds split from a line of reptilian evolution separate from, and long after, the reptilian lineage that led to mammals. However, it is instructive to consider bird blood flow patterns. In birds, the blood flow out of the heart is simplified to two vessels; a single systemic arch (from arch #4) and a single pulmonary arch (from arch #6). Birds retain the right systemic arch. The left side has been lost. Birds are derived from diapsid reptiles (diapsid = with two temporal fenestrae or "windows" in the skull) and probably came from the saurischian order of dinosaurs. The closest living retile to birds are the crocodiles. The use of only the right aortic arch in birds is perhaps homologous to the dominance of the right side in extant crocodiles. In birds, the carotid duct and ductus arteriosus are closed. The subclavian arteries in birds branch from the internal carotids (arch #3) whereas in extant reptiles they come off the systemic arches (#4).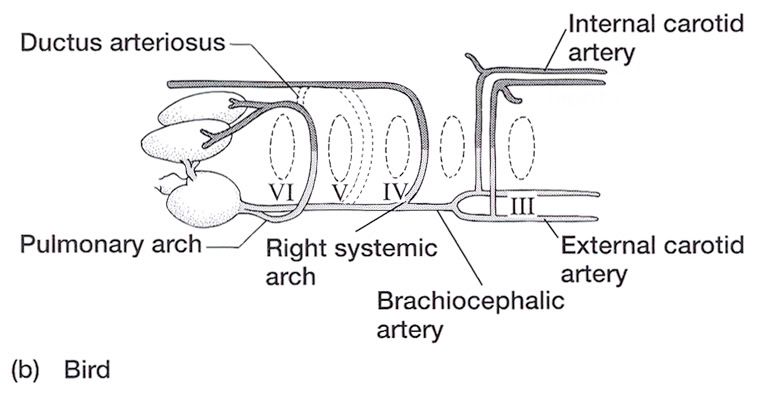 Mammals Mammals also simplify the blood circulation leaving the heart by dropping one side of the systemic arch. In contrast to birds, mammals retain the left systemic arch but lose the right side . A remnant of the ride side is retained to form the right subclavian artery. A single systemic arch is arguably more efficient than two. Both birds and mammals achieved homeothermy through ever more efficient metabolisms and converged on a single arch for systemic supply to tissues caudal to the heart. It may be arbitrary which side is retained and which is lost. Not much is known of the blood vessels of the extinct synapsid reptiles (synapsid = a single temporal fenestra in the skull) that gave rise to mammals. Blood vessels do not typically leave fossilized remains.Mammals otherwise show the same pattern of blood flow as frogs, reptiles and birds . The carotid arteries that supply the head are formed by the third aortic arch. The aortic arch is formed by arch #4. It leads cranially toward the ancestral gills in the pharynx, then bends sharply to the left as if to supply the gills on the left side of the pharynx (the right side now lost) and continues caudally to supply body tissues caudal to the heart. The single pulmonary artery, derived from aortic arch #6 leads from the right ventricle to the lungs. 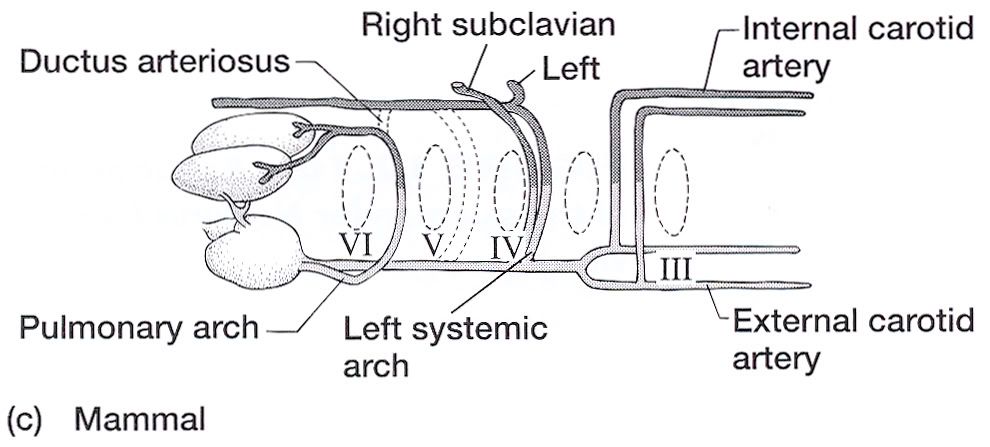 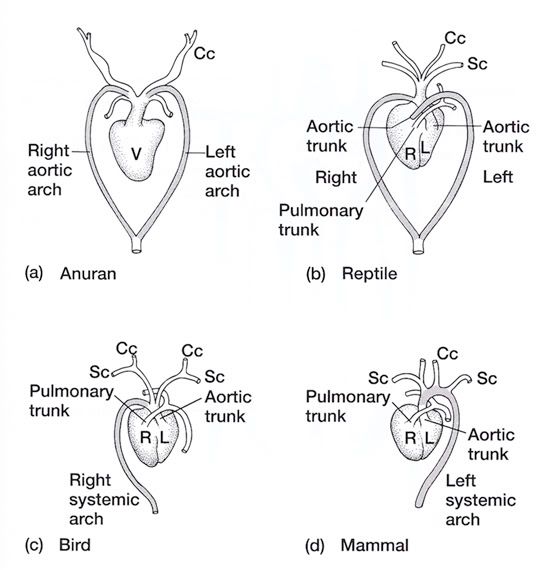 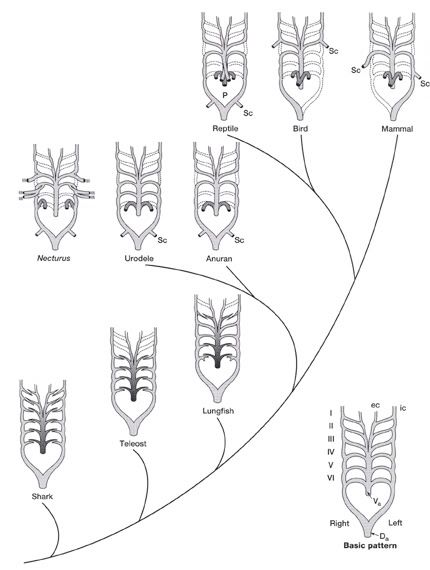
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#86
Thursday, May 21, 2009
|
|||
|
|||
|
Stages in chick embryo development One of the greatest miracles of nature is the transformation of the egg into the chick. A chick emerges after a brief three weeks of incubation. The complexity of the development cannot be understood without training in embryology.When the egg is laid, some embryonic development has occurred and usually stops until proper cell environmental conditions are established for incubation to resume. At first, all the cells are alike, but as the embryo develops, cell differences are observed. Some cells may become vital organs; others become a wing or leg. Soon after incubation begins, a pointed thickened layer of cells becomes visible in the caudal or tail end of the embryo. This pointed area is the primitive streak, and is the longitudinal axis of the embryo. From the primitive streak, the head and backbone of the embryo develop. A precursor of the digestive tract forms; blood islands appear and will develop later into the vascular or blood system; and the eye begins. On the second day of incubation, the blood islands begin linking and form a vascular system, while the heart is being formed elsewhere. By the 44th hour of incubation, the heart and vascular systems join, and the heart begins beating. Two distinct circulatory systems are established, an embryonic system for the embryo and a vitelline system extending into the egg. At the end of the third day of incubation, the beak begins developing and limb buds for the wings and legs are seen. Torsion and flexion continue through the fourth day. The chick's entire body turns 90o and lies down with its left side on the yolk. The head and tail come close together so the embryo forms a "C" shape. The mouth, tongue, and nasal pits develop as parts of the digestive and respiratory systems. The heart continues to enlarge even though it has not been enclosed within the body. It is seen beating if the egg is opened carefully. The other internal organs continue to develop. By the end of the fourth day of incubation, the embryo has all organs needed to sustain life after hatching, and most of the embryo's parts can be identified. The chick embryo cannot, however, be distinguished from that of mammals. The embryo grows and develops rapidly. By the seventh day, digits appear on the wings and feet, the heart is completely enclosed in the thoracic cavity, and the embryo looks more like a bird. After the tenth day of incubation, feathers and feather tracts are visible, and the beak hardens. On the fourteenth day, the claws are forming and the embryo is moving into position for hatching. After twenty days, the chick is in the hatching position, the beak has pierced the air cell, and pulmonary respiration has begun. After 21 days of incubation, the chick finally begins its escape from the shell. The chick begins by pushing its beak through the air cell. The allantois, which has served as its lungs, begins to dry up as the chick uses its own lungs. The chick continues to push its head outward. The sharp horny structure on the upper beak (egg tooth) and the muscle on the back of the neck help cut the shell. The chick rests, changes position, and keeps cutting until its head falls free of the opened shell. It then kicks free of the bottom portion of the shell. The chick is exhausted and rests while the navel openings heal and its down dries. Gradually, it regains strength and walks. The incubation and hatching is complete. The horny cap will fall off the beak within days after the chick hatches. EVENTS IN EMBRYONIC DEVELOPMENT Before Egg Laying: Fertilization Division and growth of living cells Segregation of cells into groups of special function (tissues) Between Laying and Incubation No growth; stage of inactive embryonic life During Incubation: First day 16 hours - first sign of resemblance to a chick embryo 18 hours - appearance of alimentary tract 20 hours - appearance of vertebral column 21 hours - beginning of nervous system 22 hours - beginning of head 24 hours - beginning of eye Second day 25 hours - beginning of heart 35 hours - beginning of ear 42 hours - heart beats Third day 60 hours - beginning of nose 62 hours - beginning of legs 64 hours - beginning of wings Fourth day - beginning of tongue Fifth day - formation of reproductive organs and differentiation of sex Sixth day - beginning of beak Eighth day - beginning of feathers Tenth day - beginning of hardening of beak Thirteenth day - appearance of scales and claws Fourteenth day - embryo gets into position suitable for breaking shell Sixteenth day - scales, claws and beak becoming firm and horny Seventeenth day - beak turns toward air cell Nineteenth day - yolk sac begins to enter body cavity Twentieth day - yolk sac completely drawn into body cavity; embryo occupies practically all the space within the egg except the air cell Twenty-first day - hatching of chick 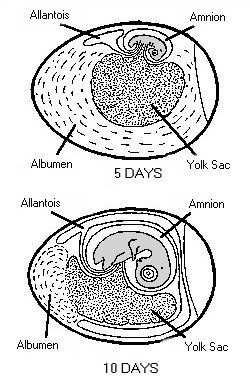 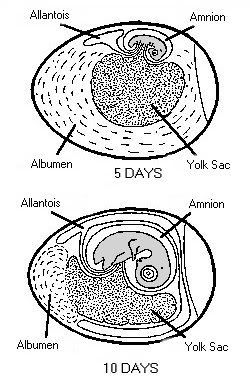 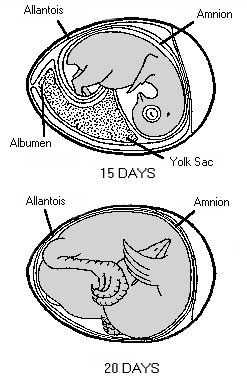 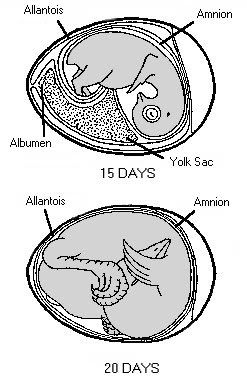
|
| The Following 3 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#87
Friday, May 22, 2009
|
|||
|
|||
|
Gastrulation in chick The images in the following presentation are not clear so I have added few images to clear the process of gastrulation. For Presentation Click Here 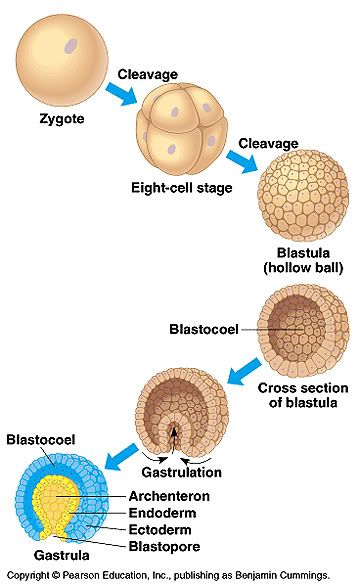 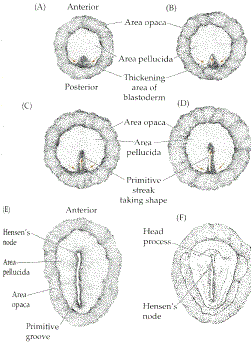 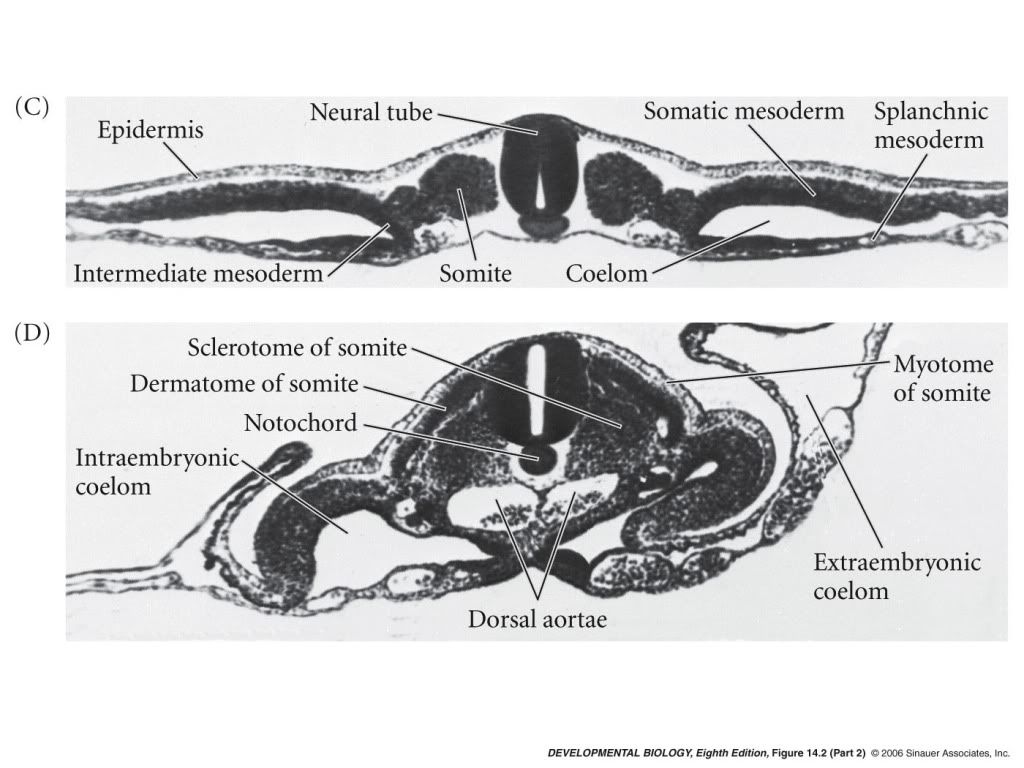 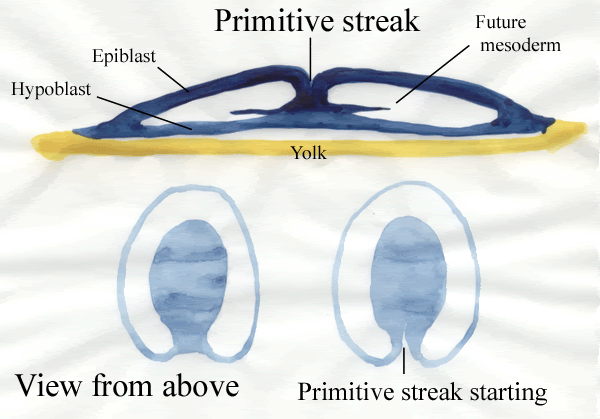 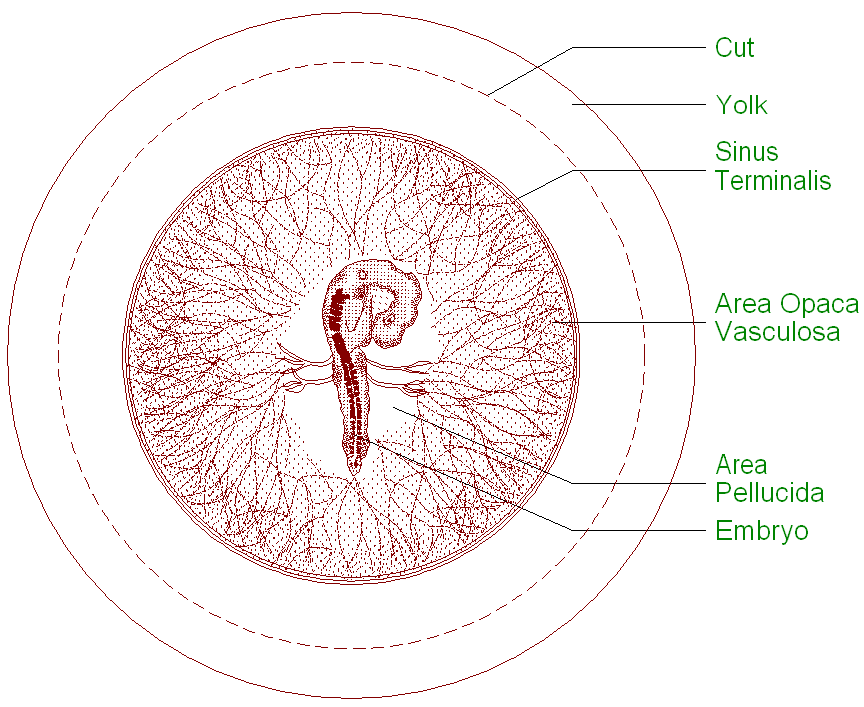 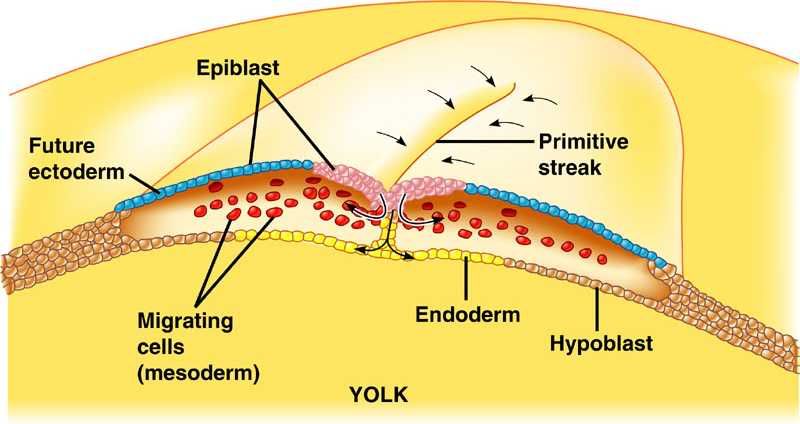
|
| The Following 3 Users Say Thank You to AFRMS For This Useful Post: | ||
dr.atifrana (Friday, May 22, 2009), Engr.Aftab (Sunday, May 24, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#88
Saturday, May 23, 2009
|
|||
|
|||
|
Comparative Kidney Structure And Function In Vertebrates. The Kidney is the principal excretory organ in a vertebrate and they are two present. In general, each consists of a mass of coelomoducts opening into a collecting duct. Kidneys are short and posterior in all vertebrates but in Fishes and Salamanders, they extend along most of the body cavity. Indeed, comparatively there are three basic types of kidneys in vertebrates.
In Reptiles, Birds, and Mammals adult kidneys (Metanephros) are non-segmental and drain wastes only from the blood . Generally, from each kidney of whatever type, a common collecting duct, the Ureter, carries the waste posteriorly. In Amphibians, Reptiles and Birds the Cloaca, to which Urinary bladder connects in Amphibians and some Reptiles. Normally, the waste or Urine is always fluid except in Reptiles and Birds, where the semisolid excretions (Uric acid) are voided as a white paste (Guano) with the faeces. In most mammals the Ureters connect directly to the bladder, while a median duct, the Urethra discharges to the exterior, passing through the Penis in the males or Vulva in females. Hence, the interrelated excretory and reproductive systems of vertebrates are commonly termed the Urogenital system. The kidney in health elaborates a urine appropriate in volume and composition to the needs of the body at all times. Physiologic anatomy of the kidney shows that each human kidney for instance, consists of an inner Medulla and an outer Cortex, the latter containing about a million minute excretory units, or Nephrons. A Nephron is made up of:
All the tubules discharge into a central cavity (Pelvis) of the kidney that connects to the Ureter. Kidney Functions in Vertebrates.
These various regulatory functions include volume control of body water, osmotic control of the extra cellular fluid, acid-base balance, body concentration of individual electrolytes, arterial blood pressure and possibly erythropoiesis The excretion of waste products such as urea, although an essential role of the kidney, is of minor biological importance compared with its contribution to homeostasis. However, according to the generally accepted theory of kidney function (by Cushy), Protein-free fluid passes from the arterioles in the Glomerulus through the Bowman capsule by ultra-filtration. This process takes place because of the pressure of the blood; the fluid in the capsule has the same percentage composition as blood Plasma minus the colloids to which the membranes are impermeable. The second step is differential reabsorption by cells of the tubules-mostly in the proximal convoluted tubule but also in the loop of Henley and the distal convoluted tubule. Reabsorption involves work with the use of oxygen and the expenditure of energy. Materials such as Glucose that are present in the blood and in the Glomerular filtrate but not in normal Urine illustrate this. Such substances are reabsorbed completely and in a direction opposite from that expected by simple diffusion. The most remarkable feature of reabsorption by the tubules is its selectivity. For example, about 1,140grams of salt (NaCl) passes from the Glomerulus's into the tubules each day ,but normally only 4 to 8 grams of this leaves the body in the Urine. The rest is reabsorbed into the blood stream. Urea, on the other hand, is constantly being removed; it is about half (30g daily) of all solids in the urine, where it is in much higher concentration than in the blood Plasma. 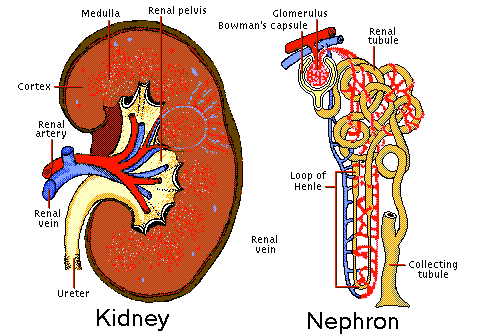 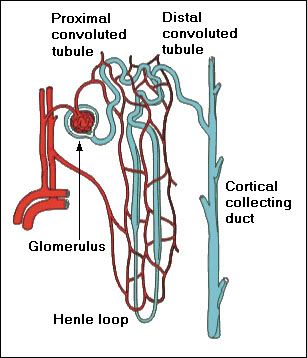 Types of kidneys in vertebrates
 Last edited by Viceroy; Saturday, May 23, 2009 at 03:31 PM. Reason: Merger
|
| The Following 4 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), prissygirl (Wednesday, September 02, 2009), Raz (Saturday, May 23, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#89
Sunday, May 24, 2009
|
|||
|
|||
|
How does the mammalian kidney produce urine? The kidney contains numerous functional units, called nephrons, which produce urine through a series of steps: glomerular filtration of the blood, tubular reabsorption of the glomerular filtrate, and tubular secretion of harmful substances. Glomerular Filtration Blood flows from the afferent arteriole into the glomerulus, a tuft of fenestrated capillaries enclosed in the Bowmans capsule. Here 15 to 25 percent of the plasmas water and solutes are filtered through a single-cell layer of the capillary walls, through a basement membrane, and into the lumen of the Bowmans capsule. The filtrate then flows into the renal tubule, to undergo tubular reabsorption. The rate of glomerular filtration depends on three factors: the hydrostatic pressure difference between the capillaries and the Bowmans capsule (due to blood pressure), the colloid osmotic pressure, which opposes filtration, and the hydraulic permeability of the three-layered tissue separating the capillaries and the lumen of the Bowmans capsule. Overall, blood pressure in the body has a major effect on the glomerular filtration rate, because the amount of blood passing through the glomerulus determines how much and how fast the fluid can be filtered. Among the dangers of very low blood pressure, therefore, is the loss of kidney function. This is a primary reason for the use of inflatable "shock suits" on the lower body in cases of extreme blood loss from trauma. By reducing blood flow to the legs, blood pressure in the trunk is kept higher, in an effort to maintain kidney function. The body through endocrine responses can regulate the glomerular filtration rate. In the case of auto regulation, increased blood pressure stretches walls of the afferent arteriole, which responds by contracting - thereby reducing fluctuation of blood pressure in the glomerulus. A drop in blood pressure brings about a decrease in the glomerular filtration rate, which, in turn, results in a decrease in sodium ions in the filtrate. (The filtrate moves through the nephrons more slowly, allowing more sodium to be reabsorbed along the way.). This lower sodium level in the filtrate is detected by the macula densa, modified cells of the wall of the distal convoluted tubule that lie adjacent to the afferent and efferent arterioles of the glomerulus. In response to the low sodium, cells of the juxtaglomerular apparatus (JGA) release rennin. This triggers a series of biochemical reactions which bring about an increase of blood pressure, and thereby an increase in GFR. This series of reactions includes an increase in angiotensin II, which helps bring blood pressure back up by (1) causing vasoconstriction in arterioles throughout much of the body, and (2) promoting increased synthesis of antidiiuretic hormone (ADH), which increases resorption of water in the collecting ducts of the kidney, thereby increasing blood volume. Angiotensin II also promotes the release of aldosterone from adrenal cortex, which promotes retention of both sodium and water, thereby helping to bring blood pressure back up. The glomerular filtration rate can also be regulated by sympathetic nerve responses, which can result in the constriction or dilation of the afferent arterioles in times of great bodily stress. Vasoconstriction increases blood pressure, and therefore GFR, whereas vasodilation decreases blood pressure and GFR. Tubular Reabsorption As the glomerular filtrate moves through the nephron, it changes composition dramatically. About 99% of the water in the original filtrate is reabsorbed, and less than 1% of the original NaCl content appears in the final urine. How does this happen? First, the filtrate moves into the proximal tubule, where about 70% of the sodium ions are reabsorbed through active transport. Water and chloride ions follow passively. Glucose and amino acids are reabsorbed here. Approximately 75% of the glomerular filtrate is reabsorbed in the proximal tubule. This is aided by the so-called "brush border" of microvilli cells which function in increasing the surface area for reabsorption. After moving through the proximal tubule, the filtrate moves on to the descending limb of the Loop of Henle. The cells in this area have no brush border, and there is no active salt transport here. The cells have a low permeability to urea and salt, but are very permeable to water. As the filtrate descends the Loop of Henle, water diffuses out because of the high salt concentration in the surrounding tissue. This is part of the urine-concentrating system of the nephron. The Loop of Henle acts as a countercurrent multiplier. For this reason, mammals living in marine or desert environments have longer loops of Henle and can conserve more water by producing more concentrated urine. The filtrate moves along the Loop of Henle to the thin segment of the ascending limb, which is highly permeable to Na+ and Cl-, and impermeable to water and urea. As the filtrate moves up the thin segment, Na+ and Cl- diffuse out because there is a higher concentration in the filtrate than in the surrounding tissues. The filtrate then moves to the medullary thick ascending limb, which is involved in the active transport of Na+ and Cl- outward from the lumen into the interstitial space. This causes the fluid reaching the distal tubule to be hypoosmotic to the interstitial fluid, and allows for the passive transport of water out of the tubule. The distal tubule functions in transporting K+, H+, and NH3 into the lumen, and Na+, Cl-, and HCO3- out. Transport of salts in this area is under endocrine control and adjusted according to osmotic conditions. The filtrate then moves into the collecting duct, which carries the fluid to the renal pelvis, to the ureters, and out of the body through the urethra. The epithelium of the collecting duct is permeable to water, but not salt or urea. The hormone ADH from the posterior pituitary gland controls the permeability o the collecting duct to water. This response causes the filtrate, which is hypoosmotic to the interstitial fluid at this point, to lose water by osmosis and therefore increase the concentration of salts and urea in the urine. At the bottom of the collecting duct, the epithelium is permeable to urea. The diffusion of some urea out of the filtrate and into the surrounding tissue helps produce the interstitial concentration gradient necessary for the diffusion of water out of the descending limb of the Loop of Henle. The urea also helps draw water out of the filtrate passing down the collecting duct, thereby enabling the kidney to excrete urine that is hypertonic to the general body fluids, a property that is important in water conservation. Tubular Secretion In several places along the nephron, substances that are not part of the initial filtrate (because the molecules are too big to be filtered through the glomerulus) are actively transported from the blood into the filtrate for elimination from the body in the urine. Some substances, such as toxins and drugs, are processed in the liver and conjugated with glucouronic acid. This marks them for removal from blood capillaries in the kidney and transport into the lumen of the nephron to become part of the filtrate. How do the kidneys regulate pH? Regulation of pH is governed by the carbon dioxide/bicarbonate buffering system in the body, which consists of three steps: CO2 + H2O <==> H2CO3 <==> HCO3- + H+ CO2 + OH- + H+ <==> HCO3- + H+ HOH <==> OH- + H+ The excretion of acid by the kidney is one of the two major factors, which influence this system (the other being the excretion of carbon dioxide by the lungs). The excretion of hydrogen ions (acid) in the urine is primarily responsible for maintaining the plasma HCO3- concentration. Mammalian urine is mildly acidic, with a pH of about 6, and contains no bicarbonate. However, the initial glomerular filtrate has a high bicarbonate concentration and a low hydrogen ion concentration. Therefore, in the process of urine formation, acid must be added to the filtrate, and bicarbonate must be removed. Therefore, the excretion of H+ and the recovery of HCO3- are both important mechanisms by which the kidneys help the body regulate pH. Special cells in the distal tubule and collecting duct, called A-type cells and B-type cells, accomplish this process. The A-type cells are acid-secreting cells that have a proton ATPase in the apical membrane and a Cl-/ HCO3- exchange system in the basolateral membrane. The cells also contain carbonic anhydrase, which hydrates carbon dioxide passing through the membrane to form protons and bicarbonate ions. The protons formed are pumped back into the lumen and can react with the bicarbonate in the filtrate to form carbon dioxide and water, which can diffuse back into the cell, and create an uptake of bicarbonate back into the blood. B-type cells, on the other hand, are base-secreting cells. They have a different form of chloride/bicarbonate exchanger in the apical membrane than the A-type cells, and secrete bicarbonate into the lumen of the tubule in exchange for chloride ions. Regulation of pH is accomplished then, by altering the activity of A and B-type cells, which determines whether bicarbonate is reabsorbed or secreted. Another mechanism used in pH regulation is the uptake of H+ by HPO4- and NH3 in the lumen to trap excess H+ in the filtrate. This occurs in order to bind H+ with something so that these protons will not move back into the epithelial cells and the blood, which would lower pH.
|
| The Following 3 Users Say Thank You to AFRMS For This Useful Post: | ||
Engr.Aftab (Sunday, May 24, 2009), prissygirl (Wednesday, September 02, 2009), sunlight (Tuesday, May 26, 2009) | ||
|
#90
Sunday, May 24, 2009
|
|||
|
|||
|
Excretion and osmoregulation The major problems that animals face with regard to osmoregulation
In most animals, the majority of cells are not in direct contact with the external environment but are bathed by an internal body fluid. Homeostatic mechanisms hamper changes in an animals body fluid, which both gives protection from harmful external environments and impedes quick exchange between intracellular compartments. The cells of the animal cannot survive much additional water gain or loss. Water continuously enters and leaves an animal cell across the plasma membrane, however, uptake and loss must balance. Animal cells swell and burst if there is a net uptake of water or shrivel and die it there is a net loss of water. Other problems associated with osmoregulation are body and environment temperatures. The enzyme activity in the body function between temperatures of 0o 40o Celsius. The way animals deal with temperature and regulating it is by way of water loss. So animals in hot environments need to limit the amount of water loss due to evaporation and respiration. The importance of water in temperature regulating leads to conflicts and compromises between physiological adaptations to environmental temperatures and osmotic stresses in terrestrial animals. The major structures involved in osmoregulation and excretion Organisms in different environments utilize different structures in osmoregulation and excretion. The major structures involved are the integument, the respiratory surface, the kidney, and the salt gland. All animals use at least one of these structures in their osmoregulatory processes. The common characteristic in structures such as gills, skin, kidneys and the integument are cells called transport epithelia, anatomically and functionally polarized cells which determine the osmoregulatory capabilities of the structure, through properties such as permeability to various solutes. The integumentfunctions in osmoregulation by acting as a barrier between the extracellular compartment and the environment to regulate water gain and loss, as well as solute flux. The permeablity of the integument to water and solutes varies from animal to animal. Respiratory surfaces such as the alveoli of the lung, and gills in aquatic animals also serve in osmoregulation and excretion. Respiratory surfaces are the chief avenues for the excretion of carbon dioxide and metabolic water, as well as other gaseous wastes, in animals. The kidney is the main organ involved in maintaining water balance and excreting harmful substances in mammals. Elasmobranchs, marine birds, and some reptiles have a structure called a salt gland to secrete NaCl from their bodies. These animals require a lower internal NaCl concentration than the surrounding seawater, which causes a concentration gradient favoring the influx of salt. Therefore, they need a way to secrete it. The solution is provided by glands in the rectum of sharks and the skulls of marine birds and reptiles which produces a concentrated salt solution for secretion. The sodium ions are removed from the blood by these glands not by filtration, but by the sodium-transport mechanism (the sodium-potassium pump). This Na+/ K+ / ATPase activity allows for the movement of NaCl from the blood across the epithelium into the lumen of the salt gland for secretion. Interestingly, the shark rectal gland, bird nasal gland, fish gill, and the thick ascending Loop of Henle in the kidney all contain salt-secreting cells that transport NaCl by the same basic mechanism. Active transport produces an increase in the chloride concentration in the cytoplasm of epithelial cells. This results in the diffusion of chloride ions out of the cell across the apical surface. The build-up of chloride ions at the apical surface attracts sodium ions to diffuse between the cells (the paracellular route). Insects have a network of Malpighian tubules extending throughout much of the body cavity and attached to the alimentary canal between the midgut and the hindgut. The secretory cells which line the walls of these long, thin tubules secrete KCl, NaCl, and phosphate from the hemolymph (blood) into the lumen of the tubule. Smaller molecules, such as water, amino acids, and sugars diffuse down their concentration gradient and into the lumen. The fluid then flows along the tubule and into the gut. As the fluid passes through the hindgut, water and valuable ions are transported back into the hemolymph, leaving behind a concentrated waste for excretion from the body. Why and how do organisms excrete metabolic wastes (particularly nitrogenous wastes)? Waste products generated in metabolic processes are often toxic, and therefore must be eliminated before they can harm the organism. The major metabolic wastes produced by animals include carbon dioxide, metabolic water, and nitrogenous wastes. Small aquatic organisms are able to get rid of wastes by simple diffusion across membranes. More complex animals with circulatory systems rely on kidneys to filter wastes out of the blood and eliminate them from the body. Carbon dioxide and metabolic water produced in respiration easily diffuse into the environment from respiratory surfaces. Nitrogenous waste excretion is more difficult, yet necessary. Elevated ammonia levels in the body can lead to convulsions, coma, and even death. This is because ammonium ions can substitute for potassium ions in ion-exchange mechanisms. Ammonia can also adversely affect metabolism and amino acid transport. An excessive amount of ammonia in the system elevates bodily pH, which causes changes in the tertiary structure of proteins, and thus cellular functions can be altered. There are three main types of nitrogenous wastes: ammonia, urea, and uric acid. The type of waste an animal excretes depends on its living environment, because nitrogenous waste excretion is accompanied by a certain amount of water loss. Ammonotelic (ammonia-excreting) animals generally live only in aquatic habitats, because ammonia is extremely toxic, and a large volume of water is required to maintain the excreted ammonia level lower than the body level. This is needed because ammonia excretion relies on passive diffusion, so a gradient is required between the organism and the environment in order for the ammonia to flow from high concentration to low concentration. Whereas most excretion of ammonia occurs across the gills of aquatic animals, mammals do excrete some ammonia in the urine. Amino groups are enzymatically transformed into glutamate, and then changed to glutamine in the liver. Glutamine can cross the kidney membranes (whereas amino acids can not). In the kidney tubules, the glutamine is deaminated to ammonia and then excreted in the urine. Although ammonia excretion is present in some forms in mammals, the major nitrogenous waste excreted is urea. Urea is less toxic than ammonia, and requires less water for elimination. Therefore, ureotelic (urea-excreting) animals are most often (but not exclusively) terrestrial. A downside to urea excretion is that urea synthesis requires energy, in the form of ATP. Vertebrates synthesize urea in the liver using the ornithine-urea cycle. Teleosts and invertebrates produce urea from uric acid via the uricolytic pathway. Birds, reptiles, and most terrestrial arthropods often are subject to very limited water availability, so even urea excretion is not possible. Therefore, these uricotelic (uric acid-excreting) animals synthesize uric acid, which requires even less water than urea for elimination. The ability to produce uric acid, which is relatively insoluble, is quite important to birds and reptiles prior to hatching. Nitrogenous wastes can be safely stored within the egg in the form of uric acid, whereas a build-up of either ammonia or urea would be deadly.
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
prissygirl (Wednesday, September 02, 2009), sunlight (Tuesday, May 26, 2009) | ||
 |
«
Previous Thread
|
Next Thread
»
|
|

 Similar Threads
Similar Threads
|
||||
| Thread | Thread Starter | Forum | Replies | Last Post |
| Very Important : How to Prepare Study Notes | Shaa-Baaz | Tips and Experience Sharing | 5 | Sunday, May 21, 2017 08:30 PM |
| Effective Study Skills | Sureshlasi | Tips and Experience Sharing | 1 | Friday, November 16, 2007 09:28 AM |
| Regarding Notes | Anonymous84 | Tips and Experience Sharing | 1 | Wednesday, August 15, 2007 06:56 PM |

