CSS Forums
Saturday, April 27, 2024
11:56 AM (GMT +5)
11:56 AM (GMT +5)





|
#51
 Wednesday, May 06, 2009
Wednesday, May 06, 2009
|
|||
|
|||
|
Phylum Arthropoda
Arthropods are multicellular, triploblastic, bilaterally symmetrical, metamerically segmented, schizocoelous, protostomous invertebrate metazoans. Cephalization of some anterior segments of the arthropods from the head is present. Externally the body is covered with a thick tough, non living, chitinous and protective cuticle, forming the exoskeleton. Exoskeleton is non-living and cannot grow. Appendages are segmental, paired, lateral and jointed and variously modified as jaws, gills and legs etc. Arthropods are triploblastic animals have true coelom. Arthropods possess separate striated muscles. More than one pair of jaw is present. Circulatory system is open, capillaries are absent and arteries open into irregular spaces called sinuses. Malpigian tubules or coelomoducts are the excretory organs that excrete ammonia, urates, amines or guanine. Nervous system is of anneidian type. Compound eye with mosaic vision is well developed. Cilia and flagella are entirely absent. Sexes are usually separate (dioecious), but a few are hermaphrodite. Sexual dimorphism is usually evident. Parthenogenesis is common in some groups. Gonads and their ducts usually paired. Fertilization is internal and development include complete, incomplete or no metamorphosis. Parental care often well marked. these are two links of PPt presentation about Arthropods Click here and check this regards 
|






| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#52
Wednesday, May 06, 2009
|
|||
|
|||
|
Phylum Arthropoda
The following is the link to detailed article on mouth parts of insects on Wikipedia check this link The link mentioned below is a comprehensive presentation on mouthparts of insects with fully labelled diagrams. click here regards
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
dr.atifrana (Wednesday, May 06, 2009), Shali (Thursday, October 08, 2009) | ||
|
#53
Wednesday, May 06, 2009
|
|||
|
|||
|
Phylum Arthropoda
Insect Metamorphosis Introduction Metamorphosis refers to a major change of form or structure during development. Insect Metamorphosis One of the most dramatic forms of metamorphosis is the change from the immature insect into the adult form. Most of the major insect orders have a typical life cycle which consists of an egg, which hatches into a larva which feeds, moults and grows larger, pupates, then emerges as an adult insect that looks very different from the larva. These insects are often called 'Holometabolous', meaning they undergo a complete (Holo = total) change (metabolous = metamorphosis or change). Those which have immature stages similar in shape to the adult minus the wings are called 'Hemimetabolous', meaning they undergo partial or incomplete (Hemi = part) change. Holometabolous (complete metamophosis) Typical holometabolous insect groups are the Coleoptera (Beetles), Lepidoptera (moths, butterflies and skippers), Hymenoptera (sawflies, wasps, ants and bees) and Diptera (flies). All these groups have a life cycle where the egg hatches into a larva (eg a caterpillar, grub, maggot) which goes through an inactive, pupa stage (eg wrapped up like a cocoon) before emerging as an adult (eg a butterfly, beetle, wasp). Hemimetabolous (incomplete metamorphosis) Typical hemimetabolous insects are the Hemiptera (Scales, Aphids, Whitefly, Cicadas, Leafhoppers and True Bugs), Orthoptera (Grasshoppers and Crickets), Mantodea (Praying Mantids), Blattodea (Cockroaches), Dermaptera (Earwigs) and Odonata (Dragonflies and Damselflies). These groups go through gradual changes as they turn into adults. Immature forms of these insects are called nymphs and these gradually increase in size and change form. As the insect grows, it sheds its skin (called moulting). After each moult, the nymph looks a bit different or a bit bigger. After a final moult, the full adult form emerges. A successful strategy Metamorphosis is one of the key elements that explains why insects are so successful. Many insects have immature stages with completely different habitats from the adults. This means that insects can often exploit valuable food resources while still being able to disperse into new habitats as winged adults. The potential for adaptation and evolution is greatly enhanced by metamorphosis. Growth and maturity There is an important feature to note regarding metamorphosis. Insects are not able to mate and reproduce until they undergo their final moult or emerge from a pupa as a winged adult. Wings do not appear until the final moult (the one exception to this is the Ephemeroptera, or Mayflies). When you see an insect with wings, it is fully grown. This means that small flies do not become larger flies, they are as big as they will get. Caterpillars, Grubs and Maggots - Holometabolous Larvae Holometabolous larvae are larvae that pupate before emerging as adult insects, and include many of the most familiar insects. Holometabolous larva in general are little more than tubular, efficient eating machines. They do not have to lay eggs, or find a mate. Apart from eating, they are mainly concerned with avoiding being eaten themselves. This means that they may have good camouflage, or hide in shelters or holes, or they may taste dreadful to any prospective predators. The major insect orders have larvae with different common names. For instance, moths, butterflies and skippers have larvae which are usually called caterpillars. Fly larvae are nearly always called maggots. Beetle larvae are often referred to as grubs. Caterpillars Moth, butterfly and skipper (Lepidoptera) caterpillars have pairs of prolegs on their abdomen in addition to the three pairs of jointed walking legs on the thorax. Prolegs differ from the usual insect legs in that they are not jointed. Each proleg has a set of tiny hooks, which are arranged in rings or series around the tip of the proleg. These are called crochets, and only occur in the insect order Lepidoptera. Although there are some caterpillar-like larvae from other insect orders, such as sawfly larvae (Order Hymenoptera, Suborder Symphyta) and leaf beetle larvae (Order Coleoptera, Family Chrysomelidae), they can be distinguished from lepidopteran larvae by the absence of prolegs with crochets. Lepidopteran larvae have chewing mouthparts, and the majority of species are adapted to eating plant material Maggots Fly larvae (Diptera) lack any segmented legs on the thorax, and are often highly specialised for living in wet environments. Very few are adapted to dry conditions. Quite a few species are internal parasites of other animals, where legs would be of no use. Unlike the larvae of Lepidoptera there is no one character that can be used to separate fly maggots from other large orders such as the Hymenoptera (Wasps, Bees, Ants, and Sawflies), as the immature stages of many species in these orders also lack segmented legs. Fly maggots live on a huge range of foods - from human flesh through to kelp on the seashore Grubs Beetle larvae (Coleoptera) are highly diverse in their shapes. The majority live in concealed habitats, such as underground, or inside trees. There are many aquatic species, and a few which resemble caterpillars and feed openly on leaves. Many retain segmented legs, although weevil grubs nearly always lack legs. Most legless beetle grubs have robust chewing mouthparts and can be distinguished from fly maggots, which often have modified mouth 'hooks'. The larvae of sawflies, wasps, bees and ants (Hymenoptera) are diverse in form. Many sawfly larvae are similar to lepidopteran caterpillars, and feed externally on plant material. The social Hymenoptera, which includes some wasps, some bees, and all ants have larvae with very few external features, as they do not have to forage for food. In these species food is brought to them by the adult nest mates. The parasitic Hymenoptera are similar in that they spend their larval period inside hosts or well-stocked nests. They do not need camouflage or legs in these habitats Nymphs, hoppers and mudeyes - Hemimetabolous insects Hemimetabolous insects do not have a pupal stage. The general appearance of the immature stages is somewhat similar to that of adults, although there may be some dramatic differences in lifestyle. Only adult insects are able to reproduce, and only adult insects have functional wings (in those species that have wings). The immature stages of these insects are generally called nymphs rather than larvae. Some have common names such as 'hoppers' (immature grasshoppers, Order Orthoptera), 'crawlers' (immature scale insects, Order Hemiptera) and 'mudeyes' (immature dragonflies, Order Odonata). Examples of hemimetabolous insects include cockroaches (Order Blattodea), crickets and grasshoppers (Order Orthoptera), stick insects (Order Phasmatodea), praying mantids (Order Mantodea), termites (Order Isoptera), dragonflies and damselflies (Order Odonata), earwigs (Order Dermaptera), sucking bugs (Order Hemiptera), wood and book lice (Order Psocoptera), and parasitic lice (Order Phithaptera). The feeding habits of hemimetabolous insects commonly mirror that of the adults, but often with a significant twist. Dragonfly nymphs are aquatic predators, but the adults are active flying insects, which hunt other flying insects. Stick insect nymphs can resemble ants, while later stage nymphs blend with the food plants. All stages of stick insects feed on plant material. The final moult between mature nymph and adult is usually accompanied by changes in colour, and in shape of the body, but there is never the dramatic difference between larvae and adult as observed in holometabolous insects.   
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#54
Thursday, May 07, 2009
|
|||
|
|||
|
Phylum Arthropoda
The importance of insects Insects represent animals commonly found in different types of the environment. They adapted to extremely harsh living conditions by developing modified, and often quite complicated mouth-parts. This helps them use all kinds of available food. It is therefore not surprising, that these widespread and numerous animals significantly affect the environment in which they live. Insects are also of great importance for the economy. Some of them are our allies whereas others are grimly fought enemies. One of the most important roles insects play in the natural word is the pollination of flower plants . Over millions of years, the evolution of flower plants and the related insects proceeded in parallel. As a result, various tools for collecting and transporting pollen have been developed, such as ventral brushes, pollen-baskets on legs or tufts of hair on other parts of the body. Some species, for instance, have unusually long tongues which help them reach the bottom of elongated flower tubes in search of nectar. Some insects pollinate flowers blooming in the daytime while others prefer flowers that open at twilight. The most important pollinators of flower plants are hymenopterans, especially wild bees, as well as lepidopterans, dipterans and coleopterans. Numerous insect species compete for food with man, causing considerable damage to crops or consuming wild plants which are also utilized by people. The chrysomelid beetles (Chrysomelidae) feed on green plant tissue. This leads to a significant decrease in the size of yield where the beetles occur in large numbers.The most famous representative of this family is the Colorado beetle (Leptinotarsa decemlineata). Other species commonly found in the Park include the red poplar leaf-beetle (Chrysomela populi), which feeds on poplars, willows and aspen, as well as one of the spotted leaf beetles, Chrysomela viginitipunctata. Insects which exercise the greatest influence on tree stands are the bark beetles (Ipidae). These miniature beetles can cause the withering of large forest areas already weakened by air pollution or severe weather conditions such as long-lasting drought. Bark beetles live mainly in wood and under the bark, the traces of boring of which are characteristic of individual species. Most changes in the spruce stands of the Wigry National Park have for years been caused by the eight-dentated bark beetle (Ips typographus). Tree stands are destroyed also by some Hymenoptera species. The mass occurrence of phytophagus hymenopterans (Symphyta) such as the pine web-spinning sawfly (Acantholyda posticalis) or the pine sawfly (Diprion pini), can severely damage the assimilation organs in trees (caterpillars feed on needles) and significantly decrease their immunity to possible attacks of other insects (e.g. one of buprestid beetles, Phaenops cyanes) or fungi, presenting as they do a considerable danger to entire tree stands. A small group of insects feeds on the blood of warm-blooded animals. This way of acquiring food is characteristic of certain dipterans such as keds (Melophagus), fowl flies (Ornithomyia), horse flies (Tabanus) and deer flies (Chrysopus), as well as various bugs (e.g. bed bug). Blood-sucking dipterans may transmit many diseases. Horse flies and deer flies may carry rabbit fever and plague germs by sucking the blood of farm animals. In spite of their arduousness as well as negative impact on the environment of man, insects play an important role in the natural world. Many species of predatory and parasitic insects significantly reduce the number of organisms which are harmful to the human economy . These insects regulate and maintain the biocoenotic balance and ensure in this way a proper functioning of the natural environment. This insect group includes, among others, all carabid species (Carabus). The larvae of most ladybird species (Coccinellidae) (e.g. two-spot lady-bird) play a similar role in nature by devouring enormous amounts of aphids, scale insects and other tiny insects. Some species of the rove beetles family (Staphylinidae) penetrate the corridors of bark beetles in search of their larvae. A similar behaviour is typical of some representatives of the chequered beetle family (Cleridae), including the ant beetle (Thanasismus formicarius) inhabiting the park. Another group of insects which plays a crucial role in different types of forest environment is ants (Formicidae). Large mound ants belonging to the Formica genus act as "orderlies" by regulating the number of other insects. In the case of the mass appearance of Lepidoptera or Diptera caterpillars feeding on plants, ants switch to these species thereby significantly reducing their number. By building their nests, ants improve the quality of the soil. Numerous chambers and corridors in the underground part of the nest have a beneficial impact on the air and water regime in the soil. Insects actively accelerate the circulation of the organic matter in the environment. The larvae of many Diptera species (e.g. bluebottle flies Caliphora and flesh-flies Sarcophaga) feed on dead plants and animals as well as on animal dung. This significant contribution leads to a faster decomposition. Carrion is a source of nourishment for numerous beetle species (e.g. burying beetles (Necrophorus) and carrion beetles Silpha). Because eggs are deposited in the carrion, the larvae feed on the animal remains. The dor beetles (Geotrupes) remove immense quantities of dung from the environment . They build deep burrows ending with chambers under an accumulation of dung where females deposit their eggs. The beetles then fill the chambers with lumps of dung providing food reserves for the developing larvae
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#55
Thursday, May 07, 2009
|
|||
|
|||
|
Phylum Arthropoda
General features » Importance to humans The crustaceans of most obvious importance to humans are the larger species, chiefly decapods. Fisheries in many parts of the world capture shrimps, prawns, spiny lobsters, and the king crab (Paralithodes) of the northern Pacific and its southern counterpart, the centolla, found off the coast of Chile. Many species of true crabssuch as the blue crab, Dungeness crab, and the stone crab, all in North America, and the edible crab of Europeare valuable sources of food. The most highly prized decapod is probably the true lobster (Homarus species), although overfishing since the early 20th century has greatly diminished the catches of both the North American and the European species. Freshwater crustaceans include crayfish and some river prawns and river crabs. Many species have only local market value. It is probable that no crustaceans are poisonous unless they have been feeding on the leaves or fruits of poisonous plants. Another crustacean, the large acorn shell (Balanus psittacus), a barnacle (order Cirripedia) measuring up to 27 centimetres (11 inches) in length, is regarded as a delicacy in South America, and a stalked barnacle (Mitella pollicipes) is eaten in parts of France and Spain. In Japan, barnacles are allowed to settle and grow on bamboo stakes, later to be scraped off and crushed for use as fertilizer. Copepods and krill are important components of most marine food webs. Planktonic (i.e., drifting) copepods, such as Calanus, and members of the order Euphausiacea (euphausiids), or krill, may be present in such great numbers that they discolour large areas of the open sea, thus indicating to fishermen where shoals of herring and mackerel are likely to be found. The water flea (Daphnia magna) and the brine shrimp (Artemia salina) are used as fish food in aquariums and fish ponds, and the larvae of the latter are widely used as food for the larvae of larger crustaceans reared in captivity. Ostracods, of which numerous fossil and subfossil species are known, are important to geologists and oil prospectors. Much damage may be done to rice paddies by burrowing crabs of various species and by the mud-eating, shrimplike Thalassina of Malaya. By undermining paddy embankments, they allow water to drain away, thus exposing the roots of the plants to the sun; if near the coast, salt water may thus be allowed to seep into the paddies. Tadpole shrimps (Triops) are often numerous in rice fields, where they stir up the fine silt in search of food, killing many of the plants. Land crabs and crayfish may damage tomato and cotton crops. Reference:Britannica
|
| The Following 3 Users Say Thank You to AFRMS For This Useful Post: | ||
dr.atifrana (Friday, May 08, 2009), Shali (Thursday, October 08, 2009) | ||
|
#56
Friday, May 08, 2009
|
|||
|
|||
|
Respiration in Insects
All insects are aerobic organisms -- they must obtain oxygen (O2) from their environment in order to survive. They use the same metabolic reactions as other animals (glycolysis, Kreb's cycle, and the electron transport system) to convert nutrients (e.g. sugars) into the chemical bond energy of ATP. During the final step of this process, oxygen atoms react with hydrogen ions to produce water, releasing energy that is captured in a phosphate bond of ATP. The respiratory system is responsible for delivering sufficient oxygen to all cells of the body and for removing carbon dioxide (CO2) that is produced as a waste product of cellular respiration. The respiratory system of insects (and many other arthropods) is separate from the circulatory system. It is a complex network of tubes (called a tracheal system) that delivers oxygen-containing air to every cell of the body. Air enters the insect's body through valve-like openings in the exoskeleton. These openings (called spiracles) are located laterally along the thorax and abdomen of most insects -- usually one pair of spiracles per body segment. Air flow is regulated by small muscles that operate one or two flap-like valves within each spiracle -- contracting to close the spiracle, or relaxing to open it. After passing through a spiracle, air enters a longitudinal tracheal trunk, eventually diffusing throughout a complex, branching network of tracheal tubes that subdivides into smaller and smaller diameters and reaches every part of the body. At the end of each tracheal branch, a special cell (the tracheole) provides a thin, moist interface for the exchange of gasses between atmospheric air and a living cell. Oxygen in the tracheal tube first dissolves in the liquid of the tracheole and then diffuses into the cytoplasm of an adjacent cell. At the same time, carbon dioxide, produced as a waste product of cellular respiration, diffuses out of the cell and, eventually, out of the body through the tracheal system. Each tracheal tube develops as an invagination of the ectoderm during embryonic development. To prevent its collapse under pressure, a thin, reinforcing "wire" of cuticle (the taenidia) winds spirally through the membranous wall. This design (similar in structure to a heater hose on an automobile or an exhaust duct on a clothes dryer) gives tracheal tubes the ability to flex and stretch without developing kinks that might restrict air flow. The absence of taenidia in certain parts of the tracheal system allows the formation of collapsible air sacs, balloon-like structures that may store a reserve of air. In dry terrestrial environments, this temporary air supply allows an insect to conserve water by closing its spiracles during periods of high evaporative stress. Aquatic insects consume the stored air while under water or use it to regulate buoyancy. During a molt, air sacs fill and enlarge as the insect breaks free of the old exoskeleton and expands a new one. Between molts, the air sacs provide room for new growth -- shrinking in volume as they are compressed by expansion of internal organs. Small insects rely almost exclusively on passive diffusion and physical activity for the movement of gasses within the tracheal system. However, larger insects may require active ventilation of the tracheal system (especially when active or under heat stress). They accomplish this by opening some spiracles and closing others while using abdominal muscles to alternately expand and contract body volume. Although these pulsating movements flush air from one end of the body to the other through the longitudinal tracheal trunks, diffusion is still important for distributing oxygen to individual cells through the network of smaller tracheal tubes. In fact, the rate of gas diffusion is regarded as one of the main limiting factors (along with weight of the exoskeleton) that prevents real insects from growing as large as the ones we see in horror movies! 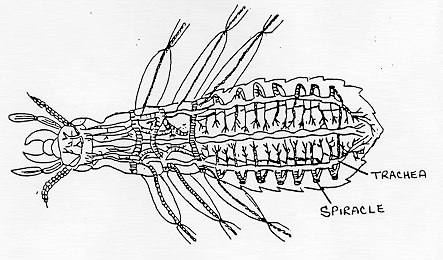 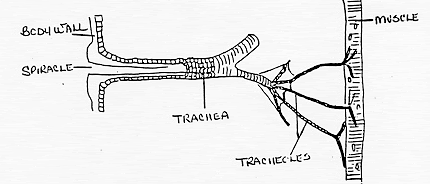 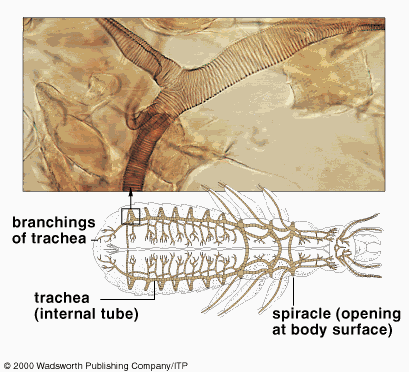 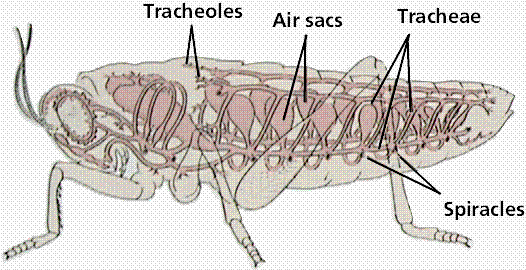
|
| The Following 2 Users Say Thank You to AFRMS For This Useful Post: | ||
dr.atifrana (Friday, May 08, 2009), Shali (Thursday, October 08, 2009) | ||
|
#57
Friday, May 08, 2009
|
|||
|
|||
|
Arthropoda appendages
Arthropods are unusual among invertebrates; they lack locomotory cilia, even as larvae. The problem that a rigid external covering imposes on movement has been solved by having the exoskeleton divided into plates over the body and through a series of cylinders around the appendages. At the junction, or joints, between the plates and cylinders the exoskeleton is thin and flexible because it lacks the exocuticle and because it is folded. The folds provide additional surface area as the joints are bent. The arthropods exoskeleton is therefore somewhat analogous to the armour encasing a medieval knight. Most arthropods move by means of their segmental appendages, and the exoskeleton and the muscles, which attach to the inside of the skeleton, act together as a lever system, as is also true in vertebrates. The external skeleton of arthropods is a highly efficient system for small animals. The exoskeleton provides a large surface area for the attachment of muscles and, in addition to functioning in support and movement, also provides protection from the external environment. The cylindrical design resists bending, and only a relatively small amount of skeletal material need be invested in thickness to prevent buckling. The external skeleton imposes limits on the maximum size of an arthropod, especially in those that live on land. The largest arthropods live in the sea, where they gain considerable support from the buoyance of seawater. On land, an excessive amount of skeleton would be required to support a large bulk and, in addition, the new soft skeleton might collapse following a molt. Appendages of arthropods have been adapted for all types of locomotionwalking, pushing, running, swimming, and burrowing. In most arthropods the legs move alternately on the two sides of the body; i.e., when one leg is in a power stroke, its mate on the opposite side of the body is in the recovery stroke (the same is true of mammals when walking). The legs in front or back are a little ahead or behind in the movement sequence. Because of the lateral position of the legs, the body of an arthropod tends to hang between them. Leg interference and trunk wobble tend to be problems in an animal with a long trunk and many legs, such as a millipede or centipede. Most arthropods have evolved more compact bodies and a smaller number of legs. The number of pairs of legs used in walking is not more than seven (crustacean pill bugs), four or five (shrimps and crabs), four (arachnids), and three (insects). This reduces the problem of mechanical interference. When a ghost crab, for example, is running rapidly across a beach or dune, only the second, third, and fourth pairs of the five pairs of legs (counting the claws) are employed. Leg interference is further reduced in most arthropods by varying limb length and placement. For example, in Scutigera, the centipede commonly seen in houses, the legs increase in length from front to back and thus pass over or under one another in stepping. The tendency for the trunk to wobble has been reduced in some centipedes by having overlapping dorsal plates and in millipedes by having pairs of segments fused to form double segments. Many arthropods are capable of walking on vertical surfaces. Some simply grip minute surface irregularities with the claws at the end of the legs. Others, such as certain spiders and flies, have an array of specialized gripping hairs at the ends of the legs. Insect wings are not segmental appendages as are the legs. The paired wings arise as lateral folds of the integument, one pair above each of the last two pairs of legs. Each wing thus consists of an upper and lower sheet of exoskeleton closely applied to each other. The two skeletal sheets are separated at various places, forming tubular supporting veins. Unlike the wings of an airplane, the wings of insects are flat plates, and lift is obtained by changing the angle at which the front margin of the wing meets the oncoming air stream. The evolution of flight is one of several adaptations that have enabled insects to become the most diverse and populous group of terrestrial animals. A burrowing habit has evolved in some insects, such as mole crickets and ants, but the largest burrowers are crustaceans. Mole crabs and box crabs are rapid burrowers in soft marine sands, and various species of mantis shrimps, mud shrimps, and snapping shrimps create elaborate burrows below the bottom surface. Crustaceans also include the largest number of arthropod tube dwellers, surpassed only by certain marine worms (polychaetes). Most of the tube-dwelling crustaceans are amphipods. Their tubes are usually composed of sand or mud particles secreted together and attached to bottom objects; there are, however, some amphipods that carry their tubes with them like a portable house. Reference:Britannica.  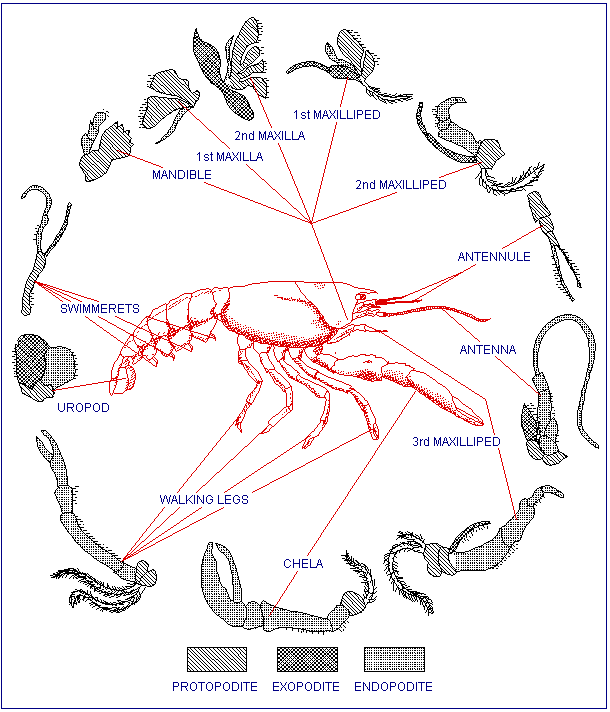 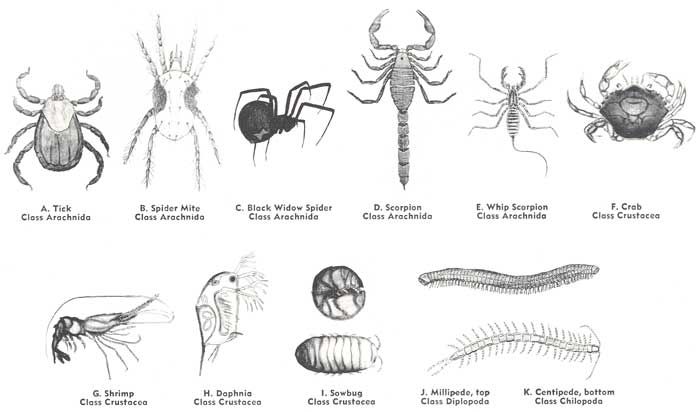 Last edited by AFRMS; Friday, May 08, 2009 at 05:15 PM. Reason: formatting
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#58
Saturday, May 09, 2009
|
|||
|
|||
|
Phylum Echinodermata
Introduction Echinoderms form a well-defined and highly-derived clade of metazoans. They have attracted much attention due to their extensive fossil record, ecological importance in the marine realm, intriguing adult morphology, unusual biomechanical properties, and experimentally manipulable embryos. The approximately 7,000 species of extant echinoderms fall into five well-defined clades: Crinoidea (sea lilies and feather stars), Ophiuroidea (basket stars and brittle stars), Asteroidea (starfishes), Echinoidea (sea urchins, sand dollars, and sea biscuits), and Holothuroidea (sea cucumbers). The phylogenetic position of the Concentricycloidea (sea daisies; 2 species), remains controversial (Baker et al. 1986; Smith 1988b; Pearse and Pearse 1994; Mooi et al. 1997). Approximately 13,000 echinoderm species are known from the fossil record. All Mesozoic and Cenozoic forms clearly fall into the five extant clades, but the Paleozoic record contains numerous distinct and often bizarre forms that have been placed into approximately 15 additional classes. Phylogenetic relationships, and in some cases status as monophyletic groups, remains unclear for the extinct classes. Unquestionable echinoderms first appear in the fossil record during the mid-Cambrian. Arkarua, a possible echinoderm, has been described from the Vendian (latest Proterozoic) (Gehling 1987). Characteristics Synapomorphies of the Echinodermata Echinoderms are among the most distinctive of all animal phyla. Inclusion in the phylum is readily diagnosable on basis of the four synapomorphies below. Most of these features are present, or can be inferred, even in the earliest fossils. Together, these synapomorphies define much of what makes the functional biology of echinoderms distinctive from that of other metazoans. Calcitic skeleton composed of many ossicles. The biomineral matrix of echinoderm skeletons is composed of calcium carbonate and several proteins. The calcite is deposited as numerous tiny crystals, but all of them lie on the same crystal axis within an ossicle. For this reason, ossicles are birefringent under polarizing light. Ossicles are not solid, but have a sponge-like microstructure called stereom that is unique to the phylum. Embryologically, echinoderm ossicles are a true endoskeleton, since they are produced by mesenchymal cells and are usually covered by epidermis. Functionally, however, the majority of ossicles act more like an exoskeleton, lying just under the epidermis and enclosing most other tissues in a flexible but tough covering. Water vascular system. The water vascular system performs many important functions in echinoderms, including locomotion, respiration, and feeding; in addition, most sensory neurons are located at the termini of podia (tubefeet) which are part of this organ system. The water vascular system may have evolved from simple tentacular systems similar to those in other deuterostome phyla, such as the tentacles of pterobranch hemichordates. However, there are many derived features of the water vascular system in echinoderms, including: an embryological origin from left mesocoel, podia arranged along branches (ambulacra), and a central circumesophageal ring. Mutable collagenous tissue. The ossicles of echinoderms are connected by ligaments composed predominantly of collagen. The material properties of this connective tissue are mutable on short timescales, under neuronal control. Ligaments are normally "locked" (rigid), but can be temporarily "unlocked" (loosened). This provides some interesting mechanical advantages, including the ability to maintain a variety of postures with no muscular effort. In holothuroids, which contain only microscopic ossicles, the entire body wall contains mutable collagenous tissue. Pentaradial body organization in adults. The adults of all extant echinoderms are radially symmetrical. A superficial bilateral organization has evolved twice, in irregular echinoids and holothuroids, but is based on an underlying five-fold organization of skeleton and most organ systems, and is clearly secondary. Higher order radial symmetry (e.g., seven-fold or nine-fold) has evolved on several occasions, and is also clearly a secondary modification. The evolutionary origins of five-fold symmetry remain obscure. Some early Paleozoic echinoderms are not radially symmetrical (e.g., carpoids and helicoplacoids), while a possible echinoderm from the Vendian (Arkarua) has five-fold radial body organization. Plesiomorphies and other features Marine habit. All extant echinoderms live in the ocean, and there is no fossil evidence of any exception to this. Within the marine realm, echinoderms occupy nearly all habitats, where they often constitute a major proportion of the biomass. Pelago-benthic life cycle. With rare exception, echinoderms are gonochoric (separate sexes) with no overt sexual dimorphism. Fertilization is almost always external. Ancestrally (and still, typically), the life cycle is complex, with a free-living larva that is planktotrophic (grazes on unicellular algae). Larvae are plesiomorphically bilaterally symmetrical, have a recurved gut and transparent ectoderm, and feed by upstream particle capture using the ciliated band. Metamorphosis is typically radical and occurs during settlement onto the benthos. Coelomate. Echinoderms form their coeloms as outpocketings from the archenteron (embryonic gut), a process called enterocoely. In most species, the coeloms are trimerous, and initially bilaterally symmetrical. The fates of the various coelomic compartments vary among echinoderms, but some features seem broadly similar and may reflect a common evolutionary origin deep within the phylum: left mesocoel gives rise to most or all of the water vascular system, and one or both somatocoels form the lining of the body cavity. Deuterostome. Like some related phyla, the blastopore (site where gastrulation begins) in echinoderm embryos becomes the larval anus; the larval mouth is a secondary opening. In some extant forms, the larval mouth is preserved as the adult mouth, while in others the entire digestive system is re-plumbed during metamorphosis and a new mouth and anus form. Simple hemal/excretory system. The hemal and excretory systems of echinoderms are linked into what Nielsen (1996) calls the "axial complex". This organ system shows similarities, and may be homologous, to those of other deuterostome phyla. In echinoderms, it is composed of: a thickened vessel (the "heart") lacking an endothelium and surrounded by a pericardium; a region where ultrafiltration occurs via podocytes; a closed circulatory system; and an opening to the external environment called the madreporite. Decentralized nervous system. The arrangment of the central nervous sytem of echinoderms is quite different from that in other deuterostomes. Radial nerves run under each of the ambulacra, and contain the cell bodies of almost all motor neurons and interneurons. A central nerve ring surrounds the gut, and is composed primarily of fiber tracks connecting the radial nerves. No known echinoderm contains anything that could be called a brain, although ganglia are present along the radial nerves in some echinoderms. Unlike most bilaterian phyla, echinoderms lack any trace of cephalization, and have no specialized sense organs. Sensory neurons are located primarily within the ectoderm of podia, and send axons to the radial nerves. 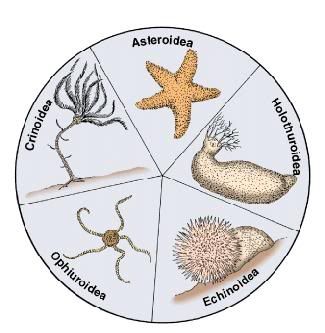
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#59
Saturday, May 09, 2009
|
|||
|
|||
|
Phylum Echinodermata
The following link is about Echinodermata,characters,classes,water vascular system. Click Here Another link to ppt presentation about Echinodermata. Check this regards
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
|
#60
Saturday, May 09, 2009
|
|||
|
|||
|
Phylum Echinodermata
Importance » Role in nature Echinoderms are efficient scavengers of decaying matter on the seafloor, and they prey upon a variety of small organisms, thereby helping to regulate their numbers. When present in large numbers, sea urchins can devastate sea-grass beds in the tropics, adversely affecting the organisms dwelling within. Sea urchins that burrow into rocks and along a shore can accelerate the erosion of shorelines. Other tropical species of sea urchins, however, control the growth of seaweeds in coral reefs, thereby permitting the corals to flourish. Removal of the sea urchins results in the overgrowth of seaweeds and the devastation of the coral reef habitat. Echinoderms can alter the structure of seafloor sediments in a variety of ways. Many sea cucumbers feed by swallowing large quantities of sediment, extracting organic matter as the sediment passes through the intestine, and ejecting the remainder. Large populations of sea cucumbers in an area can turn over vast quantities of surface sediments and can greatly alter the physical and chemical composition of the sediments. Burrowing starfish, sand dollars, and heart urchins disturb surface and subsurface sediments, sometimes to depths of 30 centimetres or more. In addition, echinoderms produce vast numbers of larvae that provide food for other planktonic organisms. Relation to human life Some of the larger species of tropical sea cucumbers, known commercially as trepang or bêche-de-mer, are dried and used in soups, particularly in Asia. Raw or cooked mature sex organs, or gonads, of sea urchins are regarded as a delicacy in some parts of the world, including parts of Europe, the Mediterranean region, Japan, and Chile. Some tropical holothurians produce a toxin, known as holothurin, which is lethal to many kinds of animals; Pacific islanders kill fish by poisoning waters with holothurian body tissues that release the toxin. Holothurin does not appear to harm human beings; in fact, the toxin has been found to reduce the rate of growth of certain types of tumours and thus may have medical significance. The eggs and spermatozoa of echinoderms, particularly those of sea urchins and starfishes, are easily obtained and have been used to conduct research in developmental biology. Indeed, echinoids have been collected in such large numbers that they have become rare or have disappeared altogether from the vicinity of several marine biologic laboratories. Starfishes that prey upon commercially usable mollusks, such as oysters, have caused extensive destruction of oyster beds. Sea urchins along the California coast have interfered with the regrowth of commercial species of seaweed by eating the young plants before they could become firmly established. The crown-of-thorns starfish, which feeds on living polyps of reef corals, has caused extensive short-term damage to coral reefs in some parts of the Pacific and Indian oceans. Reference:Britannica
|
| The Following User Says Thank You to AFRMS For This Useful Post: | ||
Shali (Thursday, October 08, 2009) | ||
 |
«
Previous Thread
|
Next Thread
»
|
|

 Similar Threads
Similar Threads
|
||||
| Thread | Thread Starter | Forum | Replies | Last Post |
| Very Important : How to Prepare Study Notes | Shaa-Baaz | Tips and Experience Sharing | 5 | Sunday, May 21, 2017 08:30 PM |
| Effective Study Skills | Sureshlasi | Tips and Experience Sharing | 1 | Friday, November 16, 2007 09:28 AM |
| Regarding Notes | Anonymous84 | Tips and Experience Sharing | 1 | Wednesday, August 15, 2007 06:56 PM |

